कान: मानव कान पर उपयोगी नोट्स (13270 शब्द)
यहाँ मानव कान पर अपने नोट्स हैं!
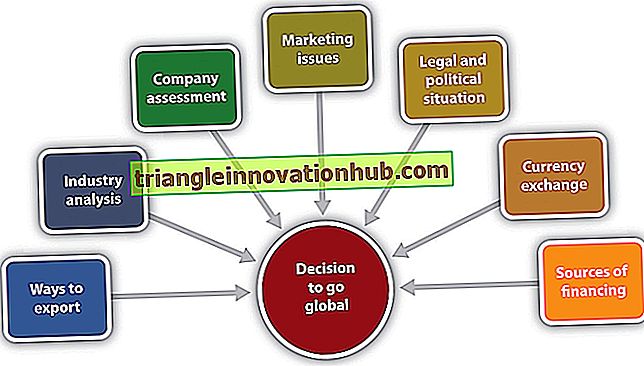
मानव कान ध्वनि आवृत्ति के प्रति संवेदनशील है 1500 या 3000 चक्र प्रति सेकंड या हर्ट्ज़ के बीच। हालांकि, युवा वयस्क कान 20 से 20, 000 हर्ट्ज तक ध्वनि आवृत्तियों के चरम के लिए उत्तरदायी हैं।

चित्र सौजन्य: lyrichearing.com/sites/default/files/assets/cutaway_ear.jpg
कान में तीन भाग होते हैं- बाहरी, मध्य और आंतरिक। बाहरी कान हवा से तंपन झिल्ली तक ध्वनि तरंगों को एकत्रित और संचालित करता है, जो सभी श्रव्य आवृत्तियों को कंपन करता है।
मध्य कान एक शरीर गुहा है जो पेट्रो टेम्पोरल के भीतर होता है, श्लेष्म झिल्ली द्वारा पंक्तिबद्ध होता है और हवा से भरा होता है जो श्रवण ट्यूब के माध्यम से नासॉफिरिन्क्स से प्राप्त होता है। मध्य कान ध्वनि कंपन के बल को तेज करता है, बिना आयाम में परिवर्तन किए, 15 से 20 गुना अधिक है, जो कि टेम्पेनिक झिल्ली की तुलना में अधिक है और कान के अस्थि-पंजर की गतिविधियों द्वारा कंपन को टेम्पेनिक झिल्ली से मेनेस्ट्रा वेस्टिबुली तक पहुंचाता है।
आंतरिक कान के पेरिलिम्फेटिक तरल पदार्थ की जड़ता को दूर करने के लिए प्रति यूनिट क्षेत्र में कंपन के बल में वृद्धि आवश्यक है। आंतरिक कान में बाहरी बोनी भूलभुलैया और आंतरिक झिल्लीदार भूलभुलैया होती है, जो पेरिलेम द्वारा अलग होती है; मेम्ब्रेनियस लेबिरिंथ एंडोलिम्फ से भरा होता है और कोर्टी के सर्पिल अंग में श्रवण के परिधीय अंगों और मैक्युला और क्राइस्ट एम्पीलारिस में संतुलन से भरा होता है।
आंतरिक कान तंत्रिका ऊर्जा में ध्वनि ऊर्जा के रूपांतरण से संबंधित है और 8 वीं कपाल तंत्रिका के कर्णावर्ती और वेस्टिबुलर विभाजन द्वारा सुनने और संतुलन की भावना को व्यक्त करता है।
विकासात्मक रूप से, आंतरिक कान पहले एक एक्टोडर्मल पुटिका, ओटोसिस्ट से प्रकट होता है; मध्य कान ट्युबो-टैंम्पिक अवकाश के एंडोडर्म से आगे विकसित होता है; बाहरी कान, एक्टोडर्मल प्रथम शाखात्मक फांक से अंतिम दिखाई देते हैं। बाहरी कान की पिन्ना ज्यादातर एक स्तनधारी विशेषता है।
बाहरी कान:
बाहरी कान में टखने और बाहरी ध्वनिक मांस होते हैं।
द टर्रिकल:
ऑरिकल या पिन्ना सिर के किनारे से एक तुरही जैसा लहराता प्रक्षेपण है और हवा से ध्वनि कंपन एकत्र करता है। बाह्य ध्वनिक मांस, अपानवायु के नीचे से अंदर तक फैले हुए हैं, जो कि तानिका झिल्ली तक होते हैं। मूल रूप से त्वचा को ढंका हुआ है, पीले लोचदार उपास्थि का एक टुकड़ा है जो बाहरी ध्वनिक मांस के कार्टिलाजीस भाग के साथ निरंतर है। हालांकि, उपास्थि कान के लोब्यूल में अनुपस्थित है, जो फाइब्रो-फैटी टिशू (चित्र। 14.1) से बना है।

टखने की विशेषताएं:
एरिकल की पार्श्व सतह अनियमित है और आगे और बाद में सामने की ओर एक समतलता प्रस्तुत करती है। यह निम्नलिखित ऊँचाइयों और अवसादों को प्रदर्शित करता है:
(ए) हेलिक्स प्रमुख परिधीय रिम बनाता है। इसका पूर्ववर्ती अंग हेलिक्स के शंकु के रूप में शंकु में उतरता है, और बाद को ऊपरी छोटे और निचले हिस्सों में विभाजित करता है। हेलिक्स के पीछे का अवरोही अंग कभी-कभी एक प्रमुख डार्विन का ट्यूबरकल प्रस्तुत करता है।
(बी) एंटीहेलिक्स एक और प्रमुख मार्जिन है जो हेलिक्स के पीछे के हिस्से के साथ समानांतर और सामने स्थित है। यह आंशिक रूप से एक गहरी अवसाद, अर्चक के शंख को घेरता है। ऊपर ट्रेस किया गया है, एंटीहेलिक्स एक अवसाद, त्रिकोणीय फोसा को घेरने वाले दो crura में विभाजित करता है। हेलिक्स और एंटीहेलिक्स के बीच संकीर्ण नाली को स्केफॉइड फोसा के रूप में जाना जाता है।
(c) शंख के अवसाद को त्रिभुज के त्रिभुजाकार फ्लैप द्वारा सामने रखा जाता है, जिसे ट्रैगस कहा जाता है। ट्रैगस के विपरीत और इंटरट्रैजिक नोच द्वारा अलग किया गया एक और त्रिकोणीय ऊंचाई, एंटीट्रैगस है।
हेलिक्स के शिखा के ऊपर शंख का छोटा हिस्सा सिम्बा पुरातत्व के रूप में जाना जाता है, जो सुपरमेटल त्रिकोण की स्थलाकृतिक स्थिति प्रदान करता है। हेलिक्स के क्रस के नीचे शंकु का बड़ा हिस्सा बाहरी ध्वनिक मांस के बाहरी उद्घाटन को प्रस्तुत करता है।
(d) फ़ाइब्रो-फैटी टिशू के एक त्वचा से ढके फ्लैप के रूप में कान का लोब्यूल एंटीट्रैगस के नीचे लटक जाता है।
Auricle की कपाल सतह प्रस्तुत करता है
प्रख्यात कॉन्टेस्टेंट, एमिनेंटिया त्रिकोणीय और अन्य, जो इसकी पार्श्व सतह के अवसादों के अनुरूप हैं।
गुदा के स्नायु:
मांसपेशियां दो सेट की होती हैं, बाहरी और आंतरिक।
बाहरी मांसपेशियों में auriculares पूर्वकाल, बेहतर और पीछे शामिल हैं। पूर्वकाल और बेहतर मांसपेशियां एपिक्रेनियल एपोन्यूरोसिस से उत्पन्न होती हैं और क्रमशः हेलिक्स की रीढ़ (ऊपरी भाग) और एरिकल की कपाल सतह के ऊपरी हिस्से में डाली जाती हैं। पश्चवर्ती मांसपेशी लौकिक हड्डी के मास्टॉयड भाग से उठती है और प्रख्यात कॉन्टेन्स्ट में डाली जाती है।
तंत्रिका आपूर्ति:
Auriculares पूर्वकाल और बेहतर चेहरे की तंत्रिका की लौकिक शाखा द्वारा आपूर्ति की जाती है, और auricularis चेहरे की तंत्रिका के पीछे auricular शाखा द्वारा खराब हो जाती है।
क्रियाएँ:
एक्सट्रिंसिक मांसपेशियां संपूर्ण रूप से टखने को हिलाती हैं। इस तरह के आंदोलन मानव जाति में महत्वहीन हैं।
आंतरिक मांसपेशियों में हेलिकॉप्टर प्रमुख और मामूली, ट्रेजिकस, एंटीट्रैजिकस, ट्रांसवर्सस एन्यूरिकुले और ओक्टिकस ऑरिकुला शामिल हैं। उनके कार्य पिना के आकार को संशोधित करना है। चूंकि अधिकांश मानव कानों में उनकी क्रियाएं न्यूनतम हैं, आंतरिक मांसपेशियों की संलग्नता दर्ज नहीं की जाती है।
तंत्रिका आपूर्ति:
टखने की पार्श्व सतह पर आंतरिक मांसपेशियों की आपूर्ति लौकिक शाखा द्वारा की जाती है, और ये कपाल की सतह पर चेहरे की तंत्रिका की पीछे की विशेष शाखा द्वारा होती हैं।
गुदा के स्नायुबंधन:
स्नायुबंधन दो सेटों के होते हैं, बाह्य और आंतरिक।
बाहरी स्नायुबंधन पूर्वकाल और पीछे हैं, जो अस्थि-पंजर को अस्थि-पंजर से जोड़ता है।
आंतरिक स्नायुबंधन, उपास्थि के उपास्थि के विभिन्न हिस्सों को जोड़ते हैं। एक मजबूत रेशेदार बैंड ट्रागस को हेलिक्स के क्रस से जोड़ता है, और सामने वाले बाहरी मांस को पूरा करता है।
त्वचा की त्वचा:
यह उपास्थि के अनुकूल है और शंकु और स्केफॉइड फोसा में कई वसामय ग्रंथियों से सुसज्जित है। कभी-कभी मोटे बाल बुजुर्ग पुरुषों में ट्रैगस, एंटीट्रैगस और इंटरट्रैजिक नोच से बाहर निकलते हैं। बालों वाली पिन्ना वाई-लिंक्ड (हॉलैंड्रिक) जीन की एक अभिव्यक्ति है।
धमनी की धमनी आपूर्ति:
(ए) बाहरी कैरोटिड धमनी की पश्च-वायुकोशीय शाखा कपाल की सतह और पार्श्व सतह के पीछे के भाग की आपूर्ति करती है।
(बी) सतही लौकिक धमनी की पूर्वकाल auricular शाखाएं, पार्श्व सतह के पूर्वकाल भाग की आपूर्ति करती हैं।
(c) पश्चकपाल धमनी की कुछ शाखाएँ कपाल की सतह के ऊपरी भाग की आपूर्ति करती हैं।
शिरापरक जल निकासी:
नसों धमनियों के साथ और सतही लौकिक और बाहरी गले की नसों में बहती हैं।
धमनी की त्वचा के नीचे धमनी-शिरापरक एनास्टोमोस कई हैं। AVA ठंडे वायुमंडलीय तापमान में फैलता है; इसलिए सर्दियों में त्वचा की दरारें टखने को प्रभावित कर सकती हैं और व्यक्ति गर्म कपड़ों के टुकड़े के साथ पिन्ना को लपेटकर ठंड के मौसम में जाने के लिए सहज महसूस करते हैं;
लसीका जल निकासी:
लिम्फैटिक में बहती है:
(ए) ट्रोगस के सामने पैरोटिड लिम्फ नोड्स;
(बी) टखने के पीछे मास्टॉयड लिम्फ नोड्स;
(c) गहरे ग्रीवा लिम्फ नोड्स का ऊपरी समूह।
संवेदी तंत्रिका आपूर्ति:
1. महान auricular तंत्रिका, अधिकांश कपाल की सतह और पार्श्व सतह के पीछे के भाग की आपूर्ति करता है जिसमें हेलिक्स, एंटीहेलिक्स और लोब्यूल शामिल हैं;
2. कम ओसीसीपिटल तंत्रिका, ऊपरी सतह को कपाल की आपूर्ति करता है;
3. ऑरिकुलो-टेम्पोरल तंत्रिका, पार्श्व सतह के पूर्ववर्ती भाग की आपूर्ति करती है, जिसमें ट्रागस, हेलिक्स का क्रस और हेलिक्स के आस-पास का क्षेत्र शामिल है;
4. वेगस तंत्रिका की विशेष शाखा, शंख और प्रख्यात कॉन्टेक्ट की आपूर्ति करती है।
यह चिकित्सकीय रूप से स्वीकार किया जाता है कि चेहरे की तंत्रिका के कुछ तंतुओं को योनि की ऑरिक्युलर शाखा के साथ होता है और शंख के क्षेत्र की आपूर्ति करता है। ऐसे तंतुओं का शरीरगत अस्तित्व अनसुलझा रहता है।
बाहरी ध्वनिक मांस:
यह शंकु के निचले भाग से तंपन झिल्ली तक फैली हुई है, और वयस्कों में लगभग 24 मिमी लंबी है (चित्र 14.2)।

कोर्स:
बाहरी मांस में एक S- आकार का वक्र होता है और इसमें तीन भाग होते हैं:
(ए) पार्स बाहरी, ऊपर की ओर निर्देशित, आगे और औसत दर्जे का;
(बी) पार्स इंटरमीडिया, ऊपर की ओर निर्देशित, पिछड़े और औसत दर्जे का;
(सी) पारस इंटर्ना, नीचे की ओर निर्देशित, आगे और औसत दर्जे का।
इसलिए, एक कान स्पेकुलम द्वारा टैंम्पेनिक झिल्ली की कल्पना करने के लिए टखने को ऊपर की ओर, पीछे की ओर और बाद में खींचा जाता है ताकि मांस को जितना संभव हो उतना सीधा बनाया जाए। उप विभाजनों:
बाहरी मांस में एक तिहाई (8 मिमी) पार्श्व में कार्टिलाजिनस हिस्सा होता है, और औसत दर्जे का दो-तिहाई (16 मिमी) में बोनी हिस्सा होता है।
उपास्थि के उपास्थि के साथ कार्टिलाजिनस भाग निरंतर होता है। मांस के ऊपरी और पीछे के भाग में उपास्थि अनुपस्थित है, और रेशेदार झिल्ली द्वारा प्रतिस्थापित किया जाता है। कभी-कभी सेंटोरिनी के तीन या अधिक विदर, कार्टिलाजिनस भाग की पूर्वकाल की दीवार को प्रभावित करते हैं।
बोनी वाला हिस्सा नीचे और सामने टेम्पोरल बोन की टिम्पेनिक प्लेट से बना होता है, और ऊपर और पीछे टेम्पोरल बोन का स्क्वैमस हिस्सा होता है। मांस के मध्य भाग के रूप में एक तंपनिक सल्कस मौजूद होता है, जहाँ तिर्यक झिल्ली को तिरछे रूप से जोड़ा जाता है, ताकि माँस का फर्श और पूर्वकाल की दीवार छत और पीछे की दीवार से अधिक लंबी हो।
कभी-कभी बोनी भाग की पूर्वकाल की दीवार एक फोरमैन, हस्क के हेटस को प्रस्तुत करती है, जो एक रेशेदार झिल्ली द्वारा बंद होती है; पूर्वकाल की दीवार की कमी टम्पेनिक प्लेट के अधूरे ओसेफिकेशन के कारण होती है। नए-जन्मे में, बोनी वाला हिस्सा विकसित नहीं होता है और हड्डी के एक tympanic अंगूठी द्वारा दर्शाया जाता है।
इसलिए, बाहरी मांस एक बच्चे में छोटा होता है और एक कान स्पेकुलम के गहरे सम्मिलन से टाइम्पेनिक झिल्ली को नुकसान हो सकता है।
कसना:
बाहरी मांस दो अवरोधों को प्रस्तुत करता है:
(ए) बोनी और कार्टिलाजिनस भाग के जंक्शन पर;
(बी) बोनी भाग में इस्थमस, जो शंकु से लगभग 2 सेमी गहरा है; यह दो अवरोधों का संकरा भाग है।
बोनी और कार्टिलाजिनस भागों के जंक्शन पर हाइपोटेरस-हीन रूप से एक ऑब्सट्यूड एंगल प्रोजेक्ट करता है। बिना मांस के विदेशी शरीर को हटाने के दौरान चोट के बिना इस कसना की सफाई एक महत्वपूर्ण कदम है।
बाहरी मांस के अस्तर उपकला:
यह त्वचा द्वारा पंक्तिबद्ध होता है, जो बाह्य रूप से त्वचा की त्वचा के साथ और आंतरिक रूप से टैंपेनिक झिल्ली की कटिस्टिक परत के साथ होता है। मांस की हड्डियों और उपास्थि के लिए त्वचा का पालन किया जाता है; इसलिए त्वचा के नीचे भड़काऊ एक्सयूडेट का संग्रह तनाव के कारण गंभीर दर्द पैदा करता है।
संशोधित कुंडलित पसीने वाली ग्रंथियां, जिन्हें सिरुंड ग्रंथियों के रूप में जाना जाता है, कार्टिलाजिनस भाग में मौजूद होती हैं; वे कान-मोम या सेरुमेन का स्राव करते हैं जो फंसे हुए पानी से अस्तर के उपकला को रोकते हैं, कीड़ों के प्रवेश का विरोध करते हैं और टाइम्पेनिक झिल्ली के प्रभावी कंपन के लिए एक गर्म आर्द्र वातावरण बनाए रखते हैं।
बाहरी मांस के संबंध:
सामने में: पैरोडिड ग्रंथि के एक हिस्से द्वारा अलग किए गए अनिवार्य के सिर;
बोनी भाग के ऊपर:
मध्य कपाल फोसा का तल और स्पर्शोन्मुख गुहा की एपिटैम्पेनिक अवकाश;
बोनी भाग के लिए बेहतर पोस्टीरियो: मास्टॉयड एंट्राम को मास्टॉयड वायु कोशिकाओं द्वारा अलग किया जाता है और केवल 1 से 2 मिमी मोटी हड्डी की पतली प्लेट होती है; मास्टॉयड एंट्रम को इस क्षेत्र (ट्रांसमीटल दृष्टिकोण) के माध्यम से शल्यचिकित्सा से खोला जा सकता है।
रक्त की आपूर्ति:
बाहरी मांस की आपूर्ति करने वाली धमनियां इससे निकली हैं:
(ए) बाहरी कैरोटिड के पीछे की विशेष शाखा;
(बी) मैक्सिलरी की गहरी auricular शाखा; तथा
(c) सतही लौकिक धमनियों की पूर्वकाल auricular शाखाएं।
शिराएं बाहरी जुगुलर और मैक्सिलरी नसों में, और बर्तनों के शिरापरक जाल में निकल जाती हैं।
तंत्रिका आपूर्ति:
(ए) छत और पूर्वकाल की दीवार, जिसे अरीकुलो-टेम्पोरल तंत्रिका द्वारा आपूर्ति की जाती है;
(बी) फर्श और पीछे की दीवार, वेगस तंत्रिका (एडरमैन की तंत्रिका) की विशेष शाखा द्वारा; यह योनि की एकमात्र त्वचीय शाखा है।
विकास:
बाहरी मसूड़ों को पहले ब्रांचियल क्लीफ्ट (हाइपो-मैंडीब्युलर क्लेफ्ट) के पृष्ठीय भाग से फफूँदयुक्त एक्टोडर्मल इनवैलिडेशन के रूप में विकसित किया जाता है।
पहले मेसिरियल फांक के बाहरी उद्घाटन के चारों ओर छह मेसोडर्मल पहाड़ियों या ट्यूबरकल से टखने को विकसित किया जाता है; तीन ट्यूबरकल कपाल की तरफ और तीन फांक के दुम की तरफ स्थित होते हैं। ये पहाड़ी एक दूसरे के साथ फ्यूज करती हैं और पिना बनाती हैं। ऐसा माना जाता है कि हेलिक्स और हेलिक्स के ट्रागस, मेन्डिबुलर ट्यूबरकल से व्युत्पन्न होते हैं, जबकि एंटी-हेलिक्स, एंटीट्रैगस और लोब्यूल को हायडॉइड ट्यूबरकल से विकसित किया जाता है।
मध्य कान:
मध्य कान या तन्य गुहा अस्थायी हड्डी के पेटू भाग के भीतर समाहित है; यह हवा से भर जाता है और श्लेष्म झिल्ली द्वारा पंक्तिबद्ध होता है। यह जन्म के समय पूर्ण वयस्क आकार ग्रहण करता है। मध्य कान श्रवण ट्यूब के माध्यम से नासो-ग्रसनी की पार्श्व दीवार के साथ सामने से संचार करता है, और पीछे मस्तूल एंट्राम के साथ एडिट्स के माध्यम से होता है।
मध्य कान बाहरी और आंतरिक कानों के बीच में बांधा जाता है, और कोरोनल सेक्शन पर यह लाल रक्त कोशिका की तरह एक द्विबीजपत्री डिस्क जैसा दिखता है, क्योंकि यह केंद्र में संकुचित होता है और परिधि (चित्र 14.2) पर बड़ा होता है, आगे एक सेक्शन के माध्यम से। पेट्रो-मास्टॉयड हड्डी की लंबी धुरी, श्रवण नलिका की एक साथ टैंपेनिक गुहा की रूपरेखा और मास्टॉयड एंट्रम एक पिस्तौल जैसा दिखता है - श्रवण ट्यूब द्वारा प्रतिनिधित्व किए जाने वाले नलिका, स्पर्शोन्मुख गुहा द्वारा शरीर, और मास्टॉयड एंट्राम द्वारा संभाल। ।
श्लेष्म अस्तर के बाहर, मध्य कान में तीन अस्थि-कलशों की एक श्रृंखला होती है- मलेसस, इनकस और स्टेपस; दो मांसपेशियां — टेंसर टायमपानी और स्टेपेडियस; और कुछ रक्त वाहिकाओं और नसों।
श्लेष एक दूसरे से सिनोवियल जोड़ों द्वारा जुड़े होते हैं और गुहा में तंपन झिल्ली से लेकर फेनेस्ट्रा वेस्टिबुली (अंडाकार खिड़की) तक फैले होते हैं। इन तरीकों से हवा में ध्वनि कंपन को बढ़ाया और आंतरिक कान तक पहुंचाया जाता है। तन्य गुहा के संरक्षण को इसके द्वारा वहन किया जाता है:
(i) श्रवण नलिका, जो कि स्पर्शरेखा झिल्ली के प्रत्येक तरफ हवा के दबाव का संतुलन बनाए रखती है;
(ii) अस्थि-पंजर के बीच की कलात्मक सतहों का आकार;
(iii) टेंसर टाइम्पनी और स्टेपेडियस का प्रतिवर्त संकुचन जो अचानक विस्फोटक ध्वनियों के दौरान क्षति को रोकता है।
उप विभाजनों:
तिर्यक गुहा में तीन भाग होते हैं:
(ए) एपिटीम्पैनम (अटारी):
टिम्पेनिक झिल्ली के ऊपर; इसमें मल्लेस का सिर, और शरीर और अगरु की छोटी प्रक्रिया शामिल है;
(बी) मेसोटाइम्पेनम:
टाइम्पेनिक झिल्ली के विपरीत; यह सबसे संकरा हिस्सा है और इसमें मल्लेस, हैंडलस की लंबी प्रक्रिया और स्टेप्स होते हैं;
(ग) हाइपोटाइम्पेनम:
झिल्ली के नीचे।
तन्य गुहा की सीमाएं:
यह मोटे तौर पर घनाकार है और छह दीवारें प्रस्तुत करता है- छत, फर्श, पूर्वकाल, पश्च, औसत दर्जे का और पार्श्व। छत फर्श की तुलना में व्यापक है, पूर्वकाल की दीवार इसकी पिछली दीवार की तुलना में संकरी है, और मध्ययुगीन और पार्श्व की दीवारें अपने उत्तलता के साथ प्रोजेक्ट करती हैं, जो कि टेनपैनियन गुहा की ओर है, जो इंटीरियर को एक ग्लासग्लास कसना बनाती है (Figs। 14.3, 14.4)


रूफ:
यह टेगमेन टिंपनी द्वारा बनाई गई है जो पेट्रो टेम्पोरल की हड्डी की एक पतली प्लेट है और टाइम्पेनिक गुहा और मध्य कपाल फोसा के बीच हस्तक्षेप करती है। बोनी प्लेट को कम और अधिक पेट्रोसेल नसों द्वारा छेद किया जाता है।
टेगमेन टाइम्पनी युवा में अप्रभावित रह सकती है, जिसके माध्यम से टाइम्पेनिक गुहा से संक्रमण सेरेब्रल मेनिन्जेस तक हो सकता है। वयस्क में, तन्य गुहा से नसें कभी-कभी छत में लगातार पेट्रोक्वैमस सिवनी के माध्यम से बेहतर पेट्रोसेल साइनस में बहती हैं, और इस तरह इंट्राक्रैनील साइनस में संक्रमण फैलती हैं।
मंज़िल:
यह टेम्पोरल बोन के पेट्रोस भाग के अंडरस्क्यूफर पर जुगुलर फोसा द्वारा बनता है, और आंतरिक जुगुलर नस के बेहतर बल्ब से संबंधित होता है। कुछ जगहों पर बोनी फोसा की कमी हो सकती है, जहां केवल श्लेष्म झिल्ली और रेशेदार ऊतक द्वारा शिरापरक गुहा को शिरा से अलग किया जाता है। कभी-कभी गौण मस्तूल वायु कोशिकाओं द्वारा फर्श को गाढ़ा और आक्रमण किया जाता है।
ग्लोसोफैरिंजियल नर्व की टिम्पेनिक शाखा फर्श को छेदती है, और कैरोटिड कैनाल के जुगुलर फोसा और निचले उद्घाटन के बीच एक एपर्चर के माध्यम से तन्य गुहा में प्रवेश करती है।
पूर्वकाल की दीवार:
यह बोनी कैरोटिड नहर की पिछली दीवार द्वारा निचले हिस्से में बंधी हुई है, जिसमें आंतरिक कैरोटिड धमनी और धमनी के चारों ओर सहानुभूति तंत्रिकाओं का एक जाल होता है। दीवार का यह हिस्सा बेहतर और अवर कैरोटिको-टायम्पेनिक वाहिकाओं और नसों द्वारा छिद्रित है।
पूर्वकाल की दीवार का ऊपरी हिस्सा दो समानांतर बोनी नहरों को प्रस्तुत करता है, एक दूसरे के ऊपर; ऊपरी नलिका टेंसर मांसपेशी के लिए, और श्रवण ट्यूब के बोनी भाग के लिए निचली नहर।
दोनों नहरें आगे, नीचे और ध्यान से गुजरती हैं, और अस्थायी अस्थि के पथरीले और स्क्वैमस भागों के जंक्शन पर खुलती हैं। एक बोनी विभाजन दो नहरों के बीच में हस्तक्षेप करता है, और तन्य गुहा की औसत दर्जे की दीवार के साथ पीछे की ओर फैलता है।
पीछे की दीवार:
यह नीचे से ऊपर व्यापक है, और निम्नलिखित विशेषताएं प्रस्तुत करता है:
(ए) दीवार का ऊपरी हिस्सा एक उद्घाटन प्रस्तुत करता है, मास्टॉयड एंट्राम के एडिटस, जिसके माध्यम से टाइम्पेनिक गुहा के एपिटैम्पेनिक भाग को मास्टॉयड एंट्रम के साथ संचार किया जाता है।
(बी) फोसा इंकुडीस एक छोटा अवसाद है जो एडिटस के निचले हिस्से के करीब है; इनगस की छोटी प्रक्रिया फोसा से एक लिगामेंट द्वारा निलंबित होती है।
(c) पीछे की दीवार के निचले हिस्से पर एक ऊर्ध्वाधर बोनी नहर का कब्जा है, जिसके माध्यम से स्टाइलो-मास्टॉयड फोरामेन तक चेहरे की तंत्रिका उतरती है।
(d) चेहरे की नहर के ऊपरी भाग से आगे की ओर एक खोखला पिरामिडनुमा प्रोजेक्ट होता है, और इसमें स्टेपेडियस मांसपेशी और इसकी तंत्रिका आपूर्ति होती है। पिरामिड के शीर्ष पर एक पिन-पॉइंट ओपनिंग है जिसके माध्यम से स्टेपेडियस का कण्डरा स्टेप्स की गर्दन के आगे से गुजरता है।
औसत दर्जे की दीवार:
औसत दर्जे की दीवार आंतरिक कान की बोनी भूलभुलैया की ओर है और निम्नलिखित विशेषताएं प्रस्तुत करती है:
(ए) प्रोमोटर आंतरिक कान के कोक्लीअ के बेसल मोड़ द्वारा निर्मित एक गोल ऊंचाई है। ग्लोसोफेरीन्जियल तंत्रिका की टेनपैनिक शाखा प्रोमोन्टोरी पर मंडराती है और टाइम्पेनिक प्लेक्सस बनाती है।
(b) फेनेस्ट्रा वेस्टिबुली (फेनस्ट्रा ओवलिस) एक रेनफॉर्मर एपर्चर है जो प्रोमोंट्री के पीछे और ऊपर स्थित है। उद्घाटन हाल के राज्य में स्टैप्स और कुंडलाकार लिगामेंट के आधार से बंद है।
(c) फेनेस्ट्रा कोक्ली (fenestra रोटुंडा) प्रोमोंटोरी के नीचे और पीछे एक छोटी खिड़की होती है, और हाल ही में एक ट्रिलमिनर माध्यमिक टिम्पेनिक झिल्ली द्वारा बंद कर दी जाती है, जो आंतरिक कान के स्केला टिंपनी से टाइमिक गुहा को अलग करती है।
(d) साइनस टिम्पनी प्रोमोनरी के पीछे एक अवसाद है और पश्च अर्धवृत्ताकार नहर के ampulla की स्थिति को इंगित करता है। प्रांतीय के सामने औसत दर्जे की दीवार का क्षेत्र कोक्लीअ के शीर्ष से निकटता से संबंधित है।
(() चेहरे की तंत्रिका नलिका का तिरछा भाग फेनस्ट्रा वेस्टिबुली के ऊपर और नीचे तब तक फैला रहता है, जब तक यह टोमेनियम की पीछे की दीवार के साथ बोनी नहर के ऊर्ध्वाधर भाग में शामिल नहीं हो जाता।
(f) प्रोसस ट्रोक्लेरिफोर्मिस एक हुक जैसी प्रक्रिया है, जो टेंसर टायपनी और श्रवण ट्यूब के लिए नहरों के बीच के बीच के विभाजन के पीछे के विस्तार से प्राप्त होती है।
टेनर टायमपानी का कण्डरा इस प्रक्रिया के आसपास बाद में मुड़ जाता है, इससे पहले कि मलीनस के हैंडल में डाला जाता है।
पार्श्व दीवार:
पार्श्व दीवार के अधिकांश का निर्माण टम्पेनिक झिल्ली की श्लेष्म कवर की औसत दर्जे की सतह से होता है, जो कि टेंपनिक गुहा के प्रति एक उत्तलता प्रस्तुत करता है; उत्तलता के अधिकतम बिंदु को ओम्बो के रूप में जाना जाता है।
श्लेष्म परत के नीचे, टेंपैनिक झिल्ली, मलले के हैंडल से लगाव देता है जो कि umbo के ऊपर से फैलता है। कॉर्डा टाइम्पनी तंत्रिका पार्स फ्लेसीडा के जंक्शन पर मैलेलस के हैंडल के औसत दर्जे की ओर को पार करती है और टाइम्पेनिक झिल्ली के पार्स टेंसा को पार करती है।
टिम्पेनिक झिल्ली के पूर्वकाल मार्जिन के करीब, पार्श्व की दीवार एक भट्ठा की तरह उद्घाटन, पेट्रोटायनामिक विदर प्रस्तुत करती है, जिसके मध्य छोर को पूर्वकाल कैनालिकस के रूप में जाना जाता है, जो कि छेरा टाइम्पनी तंत्रिका के पूर्वकाल तंत्रिका और पूर्वकाल स्नायु को प्रेषित करता है। मेलियस।
कॉर्डा टाइम्पनी तंत्रिका के लिए पीछे के कैनालिकुलस पोस्टीरियर और पार्श्व दीवारों के जंक्शन पर टिम्पेनिक झिल्ली के पीछे स्थित है।
पार्श्व की दीवार के एपिटैम्पेनिक भाग का गठन अस्थायी हड्डी के स्क्वैमस भाग द्वारा किया जाता है। इसमें सिर और मलद्वार की पूर्वकाल प्रक्रिया, शरीर और धूप की छोटी प्रक्रिया शामिल है।
कान का पर्दा:
टाइम्पेनिक झिल्ली या कान-ड्रम एक अंडाकार, अर्ध-पारदर्शी, नाशपाती ग्रे ट्रिलमिनार झिल्ली है और बाहरी ध्वनिक मांस से टाइम्पेनिक गुहा को अलग करता है। इसका अधिकतम व्यास 9-10 मिमी और न्यूनतम 8-9 मिमी (छवि 14.5) है।

पद:
झिल्ली को बाहरी रूप से बाहरी ध्वनिक मांस के फर्श के साथ लगभग 55 ° का तीव्र कोण बनाते हुए रखा जाता है। नवजात शिशु में, झिल्ली लगभग क्षैतिज होती है; इसलिए एक बच्चा वयस्क से बेहतर 'शोर की आवाज' का सामना कर सकता है।
झिल्ली की परिधि एक फ़ाइब्रो-कार्टिलाजिनस रिंग से बनी होती है, जो बाहरी ध्वनिक मांस के तल पर टायम्पेनिक प्लेट के एक सल्फ से जुड़ी होती है। सल्फास ऊपर की कमी है, जिसमें से दो छोरों से पूर्वकाल और पीछे के मर्लेओलर सिलवटों में तंपन झिल्ली के ऊपरी हिस्से में मलीस की पार्श्व प्रक्रिया में परिवर्तित हो जाती है।
झिल्ली के उपखंड:
मैलेलेओलर सिलवटों को टेंपैनिक झिल्ली को दो भागों में विभाजित करते हैं:
(ए) पारस फ्लेसीडा एक छोटा, त्रिभुजाकार शिथिल क्षेत्र है जो मैलोलर सिलवटों के ऊपर है; कभी-कभी यह एक छोटा छिद्र प्रस्तुत करता है।
(b) पार्स टेंसा बाकी झिल्ली पर कब्जा कर लेता है, जो मैलेलस के हैंडल के लगाव से और हैंडल से मध्यवर्ती परत के विकीर्ण तंतुओं के फैलाव द्वारा तना हुआ होता है।
झिल्ली की सतहों:
पार्श्व सतह अवतल और आगे की ओर निर्देशित है, आगे और बाद में, "हालांकि एक अग्रिम के रूप में जमीन से परिलक्षित ध्वनियों को पकड़ने के लिए" (ग्रांट, जेसीबी)। नतीजतन, पूर्वकाल की दीवार और बाहरी मांस के फर्श पीछे की दीवार और छत से अधिक लंबे होते हैं।
झिल्ली की औसत दर्जे की सतह उत्तल होती है और स्पर्शरेखा गुहा की ओर उभार लेती है; उत्तलता के अधिकतम बिंदु को ओम्बो के रूप में जाना जाता है। इस सतह को मैलेलस के संभाल का लगाव प्राप्त होता है, जो रेशेदार और श्लेष्म परतों के बीच हस्तक्षेप करता है और झिल्ली के केंद्र तक नीचे और कुछ पीछे की ओर झुकाव के साथ ऊपर से फैलता है।
यहाँ कोरलस टिम्पनी तंत्रिका द्वारा मललीस के हैंडल को औसतन पार किया जाता है जो पार्स फ्लेसीडा और पार्स टेंसा के जंक्शन पर झिल्ली की रेशेदार और श्लेष्म परतों के बीच आगे बढ़ता है।
झिल्ली की संरचना:
बाहर से अंदर की ओर, तंपन झिल्ली में तीन स्ट्रैट्स होते हैं:
1. बाहरी क्यूटिकल परत को वायुहीन, केराटिनाइज्ड स्तरीकृत स्क्वैमस एपिथेलियम द्वारा पंक्तिबद्ध किया जाता है और बाहरी ध्वनिक मांस की त्वचा के साथ निरंतर होता है। त्वचीय परत त्वचीय पेपिला से रहित है।
2. मध्यवर्ती रेशेदार परत में बाहरी विकिरण और आंतरिक परिपत्र फाइबर होते हैं। विकिरणशील तंतु मलद्वार की परिधि से परिधि तक जाते हैं; वृत्ताकार फाइबर परिधि में प्रचुर मात्रा में होते हैं और केंद्र में बिखरे हुए होते हैं। पार्स फ्लेसीसिडा में, रेशेदार स्ट्रैटम को ढीले संयोजी ऊतक द्वारा बदल दिया जाता है।
3. आंतरिक श्लेष्म परत झिल्ली के ऊपरी भाग में सिलिअटेड कोशिकाओं के पैच के साथ सरल स्तंभ या स्क्वैमस एपिथेलियम द्वारा पंक्तिबद्ध होती है।
श्लेष्मा झिल्ली तीन अवकाश प्रस्तुत करता है:
(ए) मर्लस के हैंडल के बीच पूर्वकाल का अवकाश और पूर्वकाल गुना जिसमें कॉर्ड टिम्पनी तंत्रिका होती है;
(बी) मैलेलस के हैंडल के बीच में पीछे की ओर और पीछे की ओर फोड़ा, जिसमें कोरडा टाइम्पनी तंत्रिका होती है;
(c) प्रूसक की प्यूस, श्लेष्म की पार्श्व प्रक्रिया से ऊपर की श्लेष्मा से भरी हुई श्लेष्मा होती है और टायपेनिक झिल्ली के म्लेच्छों और पार्स फ्लेसीसिडा की गर्दन के बीच में हस्तक्षेप करती है।
धमनी आपूर्ति:
टिम्पेनिक झिल्ली की आपूर्ति निम्न धमनियों द्वारा की जाती है:
1. मैक्सिलरी धमनी की गहरी auricular शाखा जो क्यूटिकल परत के नीचे स्थित होती है;
2. पीछे की auri-cular, और मैक्सिलरी धमनियों की पूर्वकाल झंझरी शाखा की स्टाइलामास्टॉइड शाखा; दोनों धमनियां श्लेष्म परत की आपूर्ति करती हैं।
शिरापरक जल निकासी:
बाहरी परत बाहरी नाल की नस में जाती है; भीतरी परत नालियों में अनुप्रस्थ साइनस और बर्तनों में जहरीला प्लेक्सस बनाती है।
तंत्रिका आपूर्ति:
क्यूटिकल परत द्वारा आपूर्ति की जाती है:
(ए) झिल्ली के ऊपरी और पूर्वकाल भाग में auriculo-लौकिक तंत्रिका;
(बी) झिल्ली के निचले और पीछे के भाग में वेगस तंत्रिका की auricular शाखा; श्लेष्म परत द्वारा आपूर्ति की जाती है;
(c) टायम्पेनिक प्लेक्सस के माध्यम से ग्लोसोफेरींजल तंत्रिका।
विकास:
Tympanic झिल्ली तीन स्रोतों से विकसित की जाती है, प्रत्येक भ्रूण की डिस्क के तीन जर्मिनल परतों का प्रतिनिधित्व करती है।
1. पहली ब्रांडीय दरार के पृष्ठीय छोर के एक्टोडर्म से क्यूटिकुलर परत विकसित होती है;
2. मध्यवर्ती तंतुमय परत आसन्न शाखाओं के मेहराब के मेसोडर्म से ली गई है;
3. भीतरी श्लेष्मा परत ट्युबो-टिम्पेनिक अवकाश के एंडोडर्म से विकसित होती है, जो पहले और दूसरे ग्रसनी थैली के पृष्ठीय सिरों के संलयन द्वारा बनाई जाती है।
कान स्पेकुलम के माध्यम से टायम्पेनिक झिल्ली की बाहरी जांच - यह निम्नलिखित विशेषताएं बताती है:
(ए) एक "शंकु का प्रकाश" झिल्ली के धमनीविस्फार चतुर्थांश से परिलक्षित होता है।
(b) झिल्ली के नीचे, केंद्र के ऊपर और आगे की ओर से फैली एक पीले रंग की लकीर के रूप में मैलेलस के हैंडल को देखा जाता है। लकीर के ऊपरी हिस्से में एक सफेद प्रमुखता म्लेच्छों की पार्श्व प्रक्रिया का प्रतिनिधित्व करती है।
(c) अर्गस की लंबी प्रक्रिया को अक्सर सफेद लकीर के रूप में देखा जाता है और मल्लेस के संभाल के ऊपरी हिस्से के समानांतर होता है।
तिर्यक झिल्ली के गहरे संबंध:
ए। एंटेरो-अवर क्वाड्रेंट - कैरोटिड कैनाल;
ख। धमनी-श्रेष्ठ क्वाड्रेंट - श्रवण ट्यूब का टेम्पेनिक उद्घाटन;
सी। पोस्टेरो-सुपीरियर क्वाड्रेंट - इनसस, स्टेप्स और फेनस्टेरा वेस्टिबुली की लंबी प्रक्रिया;
घ। पोस्टेरो-अवर क्वाड्रेंट-प्रोमोनरी और फेनेस्ट्रा कोक्ली।
तन्य गुहा की सामग्री:
श्लेष्म अस्तर के बाहर, तन्य गुहा में निम्नलिखित शामिल हैं:
(ए) तीन अस्थि-मलबे, धूप और स्टेपस;
(बी) दो मांसपेशियां- टेंसर टाइम्पनी और स्टेपेडियस;
(c) धमनियों के छह सेट;
(d) नसों के चार सेट।
कान के छिद्र:
मेलियस:
इसमें एक गोलाकार सिर, गर्दन और तीन प्रक्रियाएँ होती हैं - हैंडल या मनुब्रियम, पार्श्व और पूर्वकाल की प्रक्रियाएँ।
सिर एपिटैम्पेनिक भाग में स्थित है और लिगामेंट द्वारा टायपैनम की छत से निलंबित है। यह एक काठी के आकार का श्लेष-अवक्षेपण बनाने वाले भस्म के साथ पीछे की ओर आर्टिकुलेट करता है; सिर की आर्टिकुलर सतह को एक कसाव द्वारा ऊपरी और निचले पहलुओं में विभाजित किया जाता है, जहां निचला पहलू एक प्रेरणा या कोगटोथ के रूप में प्रोजेक्ट करता है।
हैंडल को नीचे की ओर, पीछे की ओर और मध्य दिशा में निर्देशित किया जाता है और इसे टैंपेनिक झिल्ली की रेशेदार परत में एम्बेड किया जाता है। संभाल के ऊपरी छोर की औसत दर्जे की सतह को टेंसर टिम्पनी के टेंडन का सम्मिलन प्राप्त होता है।
पूर्वकाल प्रक्रिया पेट्रोएंटामेनिक विदर के माध्यम से स्पैनॉइड हड्डी की रीढ़ की हड्डी के पूर्वकाल लिगामेंट से जुड़ी होती है।
पार्श्व प्रक्रिया हैंडल की जड़ से निकलती है और पूर्वकाल और पीछे के मैलेलेओलर सिलवटों के माध्यम से टिम्पेनिक सल्कस के ऊपरी भाग से जुड़ी होती है।
मैलेलस को पहले ब्रांचियल आर्क से विकसित किया गया है।
स्थूणक:
यह एक प्रीमियर दांत के आकार जैसा दिखता है, और इसमें एक शरीर, एक छोटी और एक लंबी प्रक्रिया होती है। शरीर एक सिनोवियल सैडल संयुक्त के गठन वाले मलेल के सिर के साथ कलाकृतियां करता है।
लघु प्रक्रिया पिछड़ी होती है और एपिगैम्पेनिक अवकाश के पश्च-अवर भाग में फोसा इन्कूडिस से लिगामेंट से जुड़ी होती है।
लंबे समय तक चलने वाली प्रक्रिया में पीछे की ओर तंपन झिल्ली में फैल जाती है और मललस के ऊपरी हिस्से के समानांतर होती है। इसकी नोक ध्यान से मुड़ती है और एक नॉब जैसा लेंटीफॉर्म नोड्यूल बनाती है, जो स्टैप्स के सिर के साथ आर्टिकुलेट होता है।
स्टेपीज़:
यह एक रकाब जैसा दिखता है, और सिर, गर्दन, पूर्वकाल और पीछे के अंग, और एक आधार या पैर-प्लेट होते हैं।
सिर एक कप के आकार का अवसाद प्रस्तुत करता है जो लेंटिफॉर्म नोड्यूल के साथ एक सिनोवियल बॉल और सॉकेट इन्कूडो-स्टेपेडियल जोड़ को बनाने के लिए प्रस्तुत करता है।
गर्दन को स्टेपेडियस मांसपेशियों के सम्मिलन के बाद प्राप्त होता है। डायवर्जिंग पूर्वकाल और पीछे के अंग एक रिनिफॉर्म बेस द्वारा जुड़े होते हैं, जो एनायुलर लिगामेंट द्वारा फेनेस्ट्रा वेस्टिबुली के मार्जिन से जुड़ा होता है।
स्टेप्स को दूसरी ब्रांचियल आर्क से विकसित किया गया है।
हड्डी बन जाना:
कान के प्रत्येक अस्थि-पंजर को आमतौर पर इंट्रा-यूटेराइन जीवन के चौथे महीने में एक केंद्र से अलग किया जाता है।
सभी श्रवण अस्थि-पंजर जन्म के समय वयस्क आकार ग्रहण करते हैं।
कान के अस्थि-पंजों की गति का तंत्र (अंजीर। 14.6, 14.7):


ध्वनि तरंगों के कंपन से तानिका झिल्ली औसत दर्जे की चलती है, और अंत में मैलेलस का संचालन उसी गति का अनुसरण करता है। इनकस के शरीर को इंकोडो-मेलेओलर संयुक्त में बंद कर दिया जाता है, और मल्लेस और इनसस दोनों एक अक्ष के चारों ओर औसतन घूमते हैं जो कि माइलस की पूर्वकाल प्रक्रिया से लेकर इनकस की छोटी प्रक्रिया तक फैली हुई है।
नतीजतन स्टेप्स का आधार फेनस्ट्रा वेस्टिबुली पर अंदर की ओर बढ़ता है, और परिणामस्वरूप असंगत पेरिल्मफेन फ़ेनेस्ट्रा कॉच पर माध्यमिक टाइम्पेनिक झिल्ली के बहिर्वाह का कारण बनता है।
स्टेप्स के आधार का निचला मार्जिन एक पूर्णक्रम के रूप में कार्य करता है और आंदोलन पैर के दोहन जैसा दिखता है, जबकि एड़ी जमीन पर टिकी हुई है। Be'ke'sy (1960) बताता है कि मध्यम ध्वनि की तीव्रता के साथ स्टेप्स एक ऊर्ध्वाधर अक्ष के चारों ओर घूमते हैं, जबकि उच्च-पच्चीकार जोर से ध्वनियों में अक्ष क्षैतिज हो जाता है।
जब इस्टैचियन कैथेटर के साथ मध्य कान को फुलाकर टम्पेनिक झिल्ली को बाद में मजबूर किया जाता है, तो मलीनस बाहर की ओर बढ़ता है, लेकिन अगरुस एक ही आंदोलन का पालन नहीं करता है, क्योंकि अंतः-मैलेओओलर संयुक्त अनलॉक हो जाता है; जिससे स्टेप्स को फेनेस्ट्रा वेस्टिबुली से फाड़ा जाने से रोका जाता है।
स्टेपस के आधार और फेनेस्ट्रा वेस्टिबुली के मार्जिन के बीच असामान्य ossification को ओटोस्क्लेरोसिस के रूप में जाना जाता है, जो प्रवाहकीय बहरापन का सबसे आम कारण है।
मध्य कान के स्नायु:
सेंसर
यह श्रवण ट्यूब के कार्टिलाजिनस भाग से और सल्कस ट्यूबे से उठता है, श्रवण ट्यूब के ऊपर एक बोनी नहर के माध्यम से पीछे की ओर से गुजरता है और तन्य गुहा की औसत दर्जे की दीवार के साथ; पेशी का कण्डरा बाद में प्रोसीकस ट्रोक्लेरी- फॉर्मिस के चारों ओर घूमता है और उसे मैलाकस के हैंडल की जड़ में डाला जाता है।
तंत्रिका आपूर्ति:
टेन्सर टाइम्पनी को बिना रिले के नाभि नाड़ीग्रन्थि के माध्यम से जबड़े की हड्डी के ट्रंक द्वारा आपूर्ति की जाती है, और तंत्रिका से औसत दर्जे के बर्तनों की मांसपेशियों तक फाइबर पहुंचाता है।
क्रिया:
यह मल्लेस के हैंडल को मध्ययुगीन रूप से खींचता है और टैंपेनिक झिल्ली को कसता है।
Stapedius:
यह एक विषम द्विभाजित पेशी है और टाइम्पेनिक गुहा के पीछे की दीवार पर पिरामिड के विभव से उत्पन्न होती है। इसकी कण्डरा एपिनेंस के शीर्ष पर एक एपर्चर से होकर गुजरती है और इसे स्टेप्स की गर्दन के पीछे की सतह में डाला जाता है।
तंत्रिका आपूर्ति:
स्टेपेडियस की आपूर्ति चेहरे की तंत्रिका द्वारा की जाती है।
क्रिया:
यह स्टैप्स को बाद में खींचता है।
दोनों टेंसर टाइम्पनी और स्टैपीडियस अनुबंध प्रतिवर्त और एक साथ, और आंतरिक ध्वनि को तेज आवाज से बचाने के लिए ध्वनि कंपन के प्रभाव को बढ़ाते हैं। स्टैपीडियस मांसपेशियों के पक्षाघात के परिणामस्वरूप हाइपरकेसिस होता है जहां एक कानाफूसी को एक शोर के रूप में दर्शाया जाता है।
मध्य कान की धमनियां:
धमनियों के छह सेटों में तन्य गुहा की आपूर्ति होती है:
1. पोस्टीरियर ऑरिक्युलर (या ओसीसीपिटल) धमनी की स्टेलोमास्टॉइड शाखा कॉर्डिया टिंपनी तंत्रिका के लिए पीछे के कैनालिकुलस के माध्यम से प्रवेश करती है;
2. मैक्सिलरी धमनी की पूर्वकाल टायम्पेनिक शाखा पेट्रोटायनामिक विदर के औसत दर्जे के अंत में प्रवेश करती है;
3. मध्य मैनिंजियल धमनी की पेट्रोसाल शाखा अधिक पेट्रोस तंत्रिका के लिए अंतराल के माध्यम से प्रवेश करती है;
4. मध्य मैनिंजियल धमनी की सुपीरियर टाइम्पेनिक शाखा टेंसर टायम्पनी पेशी के लिए नहर में प्रवेश करती है;
5. श्रवण ट्यूब के साथ आरोही ग्रसनी धमनी और धमनी नलिका की धमनी से शाखाएं;
6. आंतरिक कैरोटिड धमनी की Tympanic शाखाएं जो कैरोटिड नहर की दीवार को छेदती हैं।
मध्य कान की नसें:
तंत्रिका गुहा में नसों के चार सेट होते हैं:
1. सुपीरियर कैरोटिड धमनी के आसपास सहानुभूति प्लेक्स से सुपीरियर और हीन कैरोटिको-टाइम्पेनिक तंत्रिका;
2. ग्लोसोफेरीन्जियल तंत्रिका की टेनपैनिक शाखा, जो तन्य गुहा के तल पर एक कैनालिकस के माध्यम से प्रवेश करती है; यह कैरोटिको-टाइम्पेनिक तंत्रिकाओं के साथ जुड़ने के बाद प्रोमोनरी के श्लेष्म अस्तर के नीचे एक तानिका जाल बनाता है।
Tympanic plexus, tympanic cavity, mastoid antrum और अधिकांश श्रवण ट्यूब के श्लेष्म अस्तर को संवेदी तंतुओं की आपूर्ति करता है। प्लेक्सस के कुछ तंतु, पेरोटिड ग्रंथि के लिए प्रीगैंग्लिओनिक सीक्रेटो-मोटर फाइबर को व्यक्त करते हैं, टेगमेन कैविटी को छेदने के बाद कम पेट्रोसाल तंत्रिका के रूप में टाइम्पेनिक गुहा छोड़ देते हैं।
3. Chorda tympani तंत्रिका श्लेष्म और तंतुमय झिल्ली की परतों के बीच आगे बढ़ती है। It is a brnach of facial nerve, and conveys taste fibres from the anterior two- thirds of the tongue (except the vallate papillae) and pre-ganglionic secreto-motor fibres for submandibular and sublingual glands.
4. Facial nerve runs in a bony canal along the medial and posterior walls of tympanic cavity, and provides three branches—greater petrosal nerve at the genicular ganglion, nerve to the stapedius muscle opposite the pyramidal eminance, and the chorda tympani nerve about 6 mm above the stylomastoid foramen.
Mucous membrane of tympanic cavity:
The tympanic mucosa is continuous with that of the auditory tube, mastoid antrum and mastoid air cells. Most of the mucous membrane is ciliated columnar except in the following areas where the cells are non- ciliated and flat: over the auditory ossicles, posterior wall, posterior part of medial wall, parts of tympanic membrane, mastoid antrum and mastoid air cells. The glands are absent in the mucosa; close to the pharyngeal opening of auditory tube, however, mucus-secreting goblet cells are abundant.
A number of vascular folds project into the tympanic cavity, making its interior honey-combed in appearance. One fold descends from the roof of the cavity to the head of malleus and body of incus; other folds invest the stapedius and tensor tympani muscles and chorda tympani nerve. Three mucous recesses are closely related to the tympanic membrane.
Clinical tests for hearing acuity:
Deafness may be due to
(a) Disease of the middle ear (conductive deafness) or
(b) A lesion in cochlear nerve or cochlea (nerve deafness).
In addition to otoscopic examination and audiometry, it can be tested as follows by applying a vibrating tuning fork:
Rinne's test:
A tuning fork is placed on the mastoid bone and, when sound is no longer heard, it is placed in front of the external ear. In a subject with normal middle ear, the vibration is still audible, since the sound conducted by way of the tympanic membrane and ossicles is greater than the sound conducted through the bone of skull. If sound is not heard in front of the ear, conductive deafness due to middle ear disease is suspected.
In a severe nerve deafness, no sound is heard in either case.
Weber's test:
A vibrating tuning fork placed over the vertex of the skull is normally heard equally in each ear. When the sound is heard better in the affected ear, the disease lies in the middle ear where fused ossicles make the bone conduction through the skull efficient. If the sound is referred to a better hearing ear, loss of hearing is due to impaired function of the auditory nerve or cochlea.
Mastoid antrum:
The mastoid or tympanic antrum is an air sinus in the petrous part of temporal bone, and grows as backward extension of the tympanic cavity. It assumes full adult size at birth; each of its walls measures about 10 mm, possessing a capacity of about 1 ml. The antrum is lined by the non-ciliated flattened cells.
सीमाएँ:
The roof is formed by the tegmen tympani, and related to the middle cranial fossa and the temporal lobe of the brain.
The, floor receives the openings of mastoid air cells which sprout from the antrum and extend into the mastoid precess. Mastoid air cells are absent at birth, and start development after two years.
The upper part of the anterior wall presents an opening, the aditus to the antrum, through which the epitympanic part of tympanic cavity communicates with the antrum. The lower part of the anterior wall is related to the descending part of the bony canal for the facial nerve.
The posterior wall is related to the sigmoid sinus separated by a thin plate of bone. Sometimes the mastoid air cells intervene between the venous sinus and the antrum.
The medial wall presents a bulging for the lateral semicircular canal, close to the aditus of the antrum. Rest of the medial wall is related with the posterior semicircular canal separated by a thin plate of bone.
The lateral wall is formed by the squamous part of temporal bone below the supra-mastoid crest. It is represented externally by the supra- meatal triangle and corresponds, on surface projection, with the cymba conchae. The thickness of the lateral wall is only 2 mm in a new born, but the thickness increases by 1 mm each year until it becomes 12 mm — 15 mm by puberty.
Mastoid air cells:
These are intercommunicating cavities which sprout from the mastoid antrum during the second year and invade the mastoid bone. The air pockets are variable in number, size and distribution, and are lined by the flattened non-ciliated cells.
According to the distribution of air cells, the mastoid process consists of the following types:
(ए) वायवीय प्रकार, जब यह वायु कोशिकाओं द्वारा पूरी तरह से अनुमति देता है;
(b) Sclerotic type, when the air cells fail to sprout and make the mastoid process a solid mass of bone;
(c) मिश्रित प्रकार, जिसमें वायु कोशिकाएँ और अस्थि मज्जा होते हैं।
The process of pneumatization may extend beyond the mastoid process and affect the following areas;—squamous part of temporal bone above the supramastoid crest, posterior part of the root of the zygoma, around the facial canal and sigmoid sinus, along the roof and floor of the tympanic cavity, around the carotic canal and along the auditory tube. These extensions are surgically important and account for various manifestations due to inflammation of air cells.
The facial nerve:
परिचय:
The facial nerve (VII cranial) consists of a motor and sensory root, the latter is also known as the nervous intermedius. It contains the following functional components:
(a) Branchio-motor fibres, to supply the muscles which are developed from the second branchial arch;
(b) Preganglionic secreto-motor fibres, for the submandibular, sublingual, lacrimal glands and glands of soft palate and nasal cavity.
(c) Taste fibres, from the anterior two-thirds of the tongue and from the soft palate;
(d) Cutaneous somato-sensory fibres, from the concha of the auricle are probably conveyed by the facial nerve.
Deep (nuclear) origin (Fig. 14.11):
1। Motor nucleus:
It is situated in the caudal part of the pons, below and in front of the abducent nucleus. The motor nucleus represents special visceral (branchial) efferent column. It presents the following nuclear sub-groups-lateral, intermediate and medial.
The lateral group supplies the buccal musculature and buccinator muscle; the intermediate group supplies orbicularis oculi, muscles of the fore head and upper face; the medial group consists of dorso- medial and ventro-medial sets—the former supplies the auricular muscles and occipito- frontalis; the latter supplies the platysma (Carpenter, 1978).
The fibres from the motor nucleus pass dorso- medially towards the caudal end of the abducent nucleus, and then run rostrally superficial to that nucleus occupying the facial colliculus of the floor of the fourth ventricle.
At the cranial end of abducent nucleus the fibres bend abruptly downwards and forwards forming an internal genu, and emerge at the lower border of the pons through the motor root.
2। Superior salivatory nucleus:
It is situated dorso-lateral to the caudal part of the motor nucleus and represents the general visceral efferent column. It gives origin to the preganglionic secreto-motor (parasympathetic) fibres which emerge through the sensory root. Therefore, the sensory root is a misnomer, because in addition to sensory fibres it conveys secreto-motor fibres.
3। Upper part of the nucleus of tractus solitaries:
It represents special visceral afferent and possibly general visceral afferent column. It receives taste sensation from the anterior two- thirds of the tongue via the chorda tympani nerve, and from the soft palate through the greater petrosal nerve.
4। Upper part of the spinal nucleus of the trigeminal nerve:
Possibly it receives cutaneous sensations from the auricle through the auricular branch of the vagus, and the cell bodies of these fibres are located in the genicular ganglion of the facial nerve.
Central connections:
(a) The cortico-nuclear fibres (pyramidal) of the opposite side control the activities of that part of the motor nucleus which supplies the muscles of the lower part of the face.
(b) That part of the motor nucleus which supplies the muscles of the forehead and eye lids, is controlled by the cortico-nuclear fibres of both sides.
Superficial origin:
Both the roots of the facial nerve are attached to the lower border of the pons between the olive and inferior cerebellar peduncle. The motor root is large and lies on the medial side of the sensory root. The vestibulo-cochlear nerve is situated just lateral to the sensory root. Sometimes the sensory root or nervous intermedius is found to emerge along with the vestibulo-cochlear nerve and gradually inclines to join the facial nerve in the internal acoustic meatus.
Course and relations:
The course of the facial nerve may be divided by the stylomastoid foramen into intracranial- intrapetrous part and extracranial part (Figs. 14.8, 14.9).


Intracranial-intrapetrous part:
From the brain stem both roots of the facial nerve, accompanied by the vestibulo-cochlear nerve, pass laterally and forwards and enter the internal acoustic meatus where the labyrinthine artery accompanies them.
In the meatus the motor root lodges in a groove on the antero-superior surface of the vestibulo-cochlear nerve, and the sensory root intervenes between them. At the bottom of the meatus the two roots of facial nerve unite to form a trunk which enters the bony facial canal.
In the bony canal at first the nerve passes laterally above the vestibule of the internal ear, and reaches the medial wall of the epitympanic part of tympanic cavity where it bends backward forming a genu (external genu). At the genu the facial nerve presents an asymmetrical swelling known as the genicular ganglion which contains the cell bodies of the pseudo-unipolar sensory neurons.
From the genu the nerve passes backward and downward lodging in the bony canal above the promontory and the fenestra vestibuli, and reaches the medial wall of the aditus to the mastoid antrum. Here the nerve is related above with a bulging of the lateral semicircular canal.
Finally the nerve passes vertically downward along the posterior wall of the tympanic cavity and leaves the temporal bone through the stylomastoid foramen.
Extracranial part:
At its exit from the stylomastoid foramen the facial nerve changes the direction, passes forward superficial to the styloid process of temporal bone and pierces the posteromedial surface of the parotid gland. Within the gland the nerve runs forward for about 1 cm superficial to the retro-mandibular vein and external carotid artery, and then divides into temporo-facial and cervico-facial trunks.
The temporo-facial turns abruptly upward and subdivides into temporal and zygomatic branches. The cervico-facial passes downward and forward and subdivides into buccal, marginal mandibular and cervical branches.
The five terminal branches radiate like the goose's foot through the anterior border of the gland and supply the facial muscles. Such branching pattern is known as the pes anserinus.
Points for special importance:
1. The facial nerve presents two genu—an internal genu within the pons around the cranial end of abducent nucleus; an external genu containing cell bodies of sensory neurons in the medial wall of the epitympanic part of tympanic cavity.
2. From the brain stem to terminal distribution, the facial nerve changes direction five times—forward and laterally, lateralward, backward, downward, and forward.
3. The stylomastoid foramen lies about 2 cm deep to the middle of the anterior border of the mastoid process. In a new born, the stylomastoid foramen lies at the surface of mastoid bone, since the mastoid process grows about the 2nd year after birth. Therefore, a skin abscess behind the auricle is incised carefully in a child in order to avoid injury of the facial nerve.
शाखाओं:
Branches of communications:
1. In the internal acoustic meatus—with the vestibulo-cochlear nerve;
2. At the genicular ganglion
(a) With the pterygo-palatine ganglion through the greater petrosal nerve, which arises from the genicular ganglion and leaves the tympanic cavity through a hiatus for that nerve in the tegment tympani.
On reaching the foramen lacerum below the trigeminal ganglion, the greater petrosal nerve joins with the deep petrosal nerve from the sympathetic plexus around the internal carotid artery and forms the nerve of the pterygoid canal; the latter appears in the pterygo-palatine fossa and joins with the pterygo-palatine ganglion.
The greater petrosal nerve conveys preganglionic secretomotor fibres to the lacrimal gland and glands of the nasal mucous membrane and the soft palate. It also conveys taste fibres from the soft palate.
The preganglionic fibres are relayed into the cells of the pterygo-palatine ganglion. The postganglionic fibres reach the lacrimal gland after passing successively through the zygomatic and lacrimal nerves, and those fibres for the glands of soft palate and nasal mucosa pass through the greater and lesser palatine nerves.
The taste fibres from the soft palate pass successively through the lesser palatine nerve, pterygo-palatine ganglion without interruption, nerve of pterygoid canal, greater petrosal nerve and end as peripheral processes of the sensory neurons of the genicular ganglion.
(b) With the otic ganglion by a branch which joins with the lesser petrosal nerve.
(c) With the sympathetic plexus around the middle meningeal artery by a branch known as the external petrosal nerve.
3. In the facial canal—with the auricular branch of vagus nerve, through which the facial nerve possibly conveys the cutaneous sensations from the concha.
4. Below the stylomastoid foramen—with the glossopharyngeal, vagus, auriculo-temporal and great auricular nerves.
5. Behind the ear—with the lesser occipital nerve.
6. In the face—with the branches of the trigeminal nerve; while the motor supply of the muscles of face is derived from the facial nerve, the proprioceptive impulses from the facial muscles are conveyed centrally by the trigeminal nerve.
7. In the neck—with the transverse cervical cutaneous nerve.
Branches of distribution
1. In the facial canal (two branches):
(a) Nerve to the stapedius:
It arises from the facial nerve opposite the pyramidal eminence and supplies the stapedius muscle. Paralysis of the stapedius produces hyperacusis.
(b) Chorda tympani nerve (Fig. 14.10):

It arises from the facial nerve about 6 mm above the stylomastoid foramen. It conveys the taste fibres from the anterior two-thirds of the tongue except vallate papillae, and the preganglionic secreto- motor fibres for submandibular and sublingual glands. The chorda tympani nerve represents the pretrematic branch of the first branchial arch.
Course and relations:
The nerve passes forward and upward accompanied by the stylomastoid artery in a separate bony canal, and appears in the posterior wall of the tympanic cavity at the posterior canaliculus for the nerve which is situated behind the tympanic membrane. The nerve passes forward between the fibrous and mucous layers of the tympanic membrane at the junctions of pars flaccida and pars tensa, and crosses the medial side of the handle of the malleus.
It leaves the tympanic cavity through the anterior canaliculus for the nerve which is situated at the medial end of the petro-tympanic fissure. Here the nerve is accompanied by the anterior ligament of the malleus and the anterior tympanic branch of maxillary artery.
The chorda tympani nerve then passes downward and forward in the infratemporal fossa under cover of the lateral pterygoid muscle, crosses the medial side of the spine of the sphenoid bone and joins at an acute angle the posterior border of the lingual nerve through which the fibres of chorda tympani are distributed. In the infra-temporal fossa, chorda tympani is related laterally with the middle meningeal artery,

2. Below the stylomastoid foramen (three branches):
(a) Posterior auricular nerve—It passes upward and backward behind the auricle, supplies the intrinsic muscles of the cranial surface of the auricle, auricularis posterior and occipital belly of occipito-frontalis.
(b) Nerve to the posterior belly of digastric;
(c) Nerve to the stylohyoid muscle; sometimes the latter two branches arise as a common trunk.
3. In the face (five terminal branches):
(a) Temporal branch passes upward and forward in front of the auricle and across the zygomatic arch, and supplies intrinsic muscles of the lateral surface of the auricle, auricularis anterior and superior muscles, upper part of orbicularis oculi, frontalis and corrugator supercilli.
(b) Zygomatic branch runs along the zygomatic arch and supplies the lower part of the orbicularis oculi.
(c) Buccal branch consists of superficial and deep parts. Superficial branches supply the procerus. Deep branches subdivide into upper and lower sets. Upper buccal passes forward above the parotid duct and supplies zygo-maticus major and minor, levator anguli oris, levator labii superioris, levator labii superioris alaeque nasi and muscles of the nose. Lower buccal passes below the parotid duct and supplies the buccinator and orbicularis oris.
(d) Marginal mandibular branch appears first in the neck, then curves upward and forward across the lower border of the mandible at the antero-inferior angle of the masseter, and appears in the face after crossing superficial to the facial artery and vein. It supplies the risorius, depressor anguli oris, depressor labii inferioris and mentalis.
(e) Cervical branch appears in the anterior triangle of neck through the apex of parotid gland and supplies the platysma.

Tracings of the constituent fibres of facial nerve (Fig. 14.11)

1. Branchio-motor fibres arising from the motor nucleus pass successively through the motor root and trunk of the facial nerve, supply stapedius muscle opposite the pyramidal eminence and the rest are distributed below the stylomastoid foramen.
2. Preganglionic secreto-motor fibres for lacrimal glands, and glands of soft palate and nasal mucosa arising from the superior salivatory nucleus (possibly lacrimatory nucleus) pass successively through the sensory root and trunk of the facial nerve, greater petrosal nerve and nerve of pterygoid canal, and the fibres are relayed into the cells of the pterygopalatine ganglion. Postganglionic fibres for the lacrimal gland pass through the zygomatic and lacrimal nerves and reach the target organ; those for the glands of soft palate and nasal mucosa descend through the greater and lesser palatine nerves.
Preganglionic fibres for the submandibular and sublingual glands arise from the superior salivatory nucleus and pass in succession through the sensory root and trunk of the facial nerve, chorda tympani and lingual nerves and terminate in the submandibular ganglion which acts as a relay station. Postganglionic fibres from the ganglion supply directly the submandibular gland and reach the sublingual gland after joining the lingual nerve.
3। Taste fibres from the anterior two-thirds of the tongue pass successively through the lingual, chorda tympani, trunk of the facial nerve and end as peripheral processes of the pseudo-unipolar sensory neurons of the genicular ganglion. Taste fibres from the soft palate pass in succession through the lesser palatine nerve, pterygo-palatine ganglion without interruption, nerve of the pterygoid canal and greater petrosal nerve and terminate as peripheral processes of the sensory neurons of the genicular ganglion.
The central processes of the genicular ganglion pass through the trunk and sensory root of facial nerve and terminate in the pons in the upper part of the nucleus of tractus solitarius. A fresh relay of fibres from the solitary nucleus crosses the middle line and ascends to the opposite side as the solitario- thalamic tract which terminates in the VPM nucleus of the thalamus. Final relay from the thalamus is projected to the lower part of the post-central gyrus (areas 3, 1, 2, ) for per-ception of taste sensations.
4। Cutaneous sensations from the concha of the auricle are possibly conveyed first by auricular branch of the vagus, and thence through the trunk of facial nerve to the genicular ganglion where the cell bodies are located; the central processes of the ganglion cells pass through the trunk and sensory root of facial nerve and make synaptic connections with the upper part of the spinal nucleus of trigeminal nerve.
In herpes zoster affecting the concha, the viral lesion is found to involve the genicular ganglion of facial nerve. The anatomical disposition of the cutaneous fibres in facial nerve awaits further confirmation.

Internal ear:
The internal ear consists of an outer bony labyrinth and an inner membranous labyrinth. The bony labyrinth includes a series of intercommunicating bony cavities within the petrous temporal, is filled with perilymph and contains the membranous labyrinth. Total length of bony labyrinth is about 18 mm, and it presents three parts from before backwards—cochlea, vestibule and three semi-circular canals (Fig. 14.13).

The membranous labyrinth consists of a closed system of intercommunicating membranous sacs and ducts within the bony labyrinth, and is filled with endolymph. It presents from before backwards three parts
(a) Cochlear duct (scala media) within the bony cochlea; it contains the spiral organ of Corti which acts as peripheral organ of hearing;
(b) Saccule and utricle within the vestibule are concerned with static equilibrium and linear acceleration by the gravity receptors of the maculae;
(c) Three semi-circular ducts within the corresponding bony canals regulate kinetic equilibrium and the angular acceleration by the rotary receptors of crista ampullaris.
Note: The internal ear, three auditory ossicles, tympanic cavity and mastoid antrum assume full adult size at birth.
Bony labyrinth:
Cochlea:
The cochlea is conical in shape and resembles the shell of a common snail. Its apex known as the cupula is directed forward and laterally to the medial wall of the tympanic cavity, and its base lies at the bottom of the internal acoustic meatus. It measures about 5 mm from apex to base, and 9 mm at the base. The cochlear consists of a central pillar, the modiolus, and a bony cochlear canal (Figs. 14.14, 14.15, 14.16).


The modiolus is conical with the apex directed to the cupula, and its base at the bottom of the meatus presents tractus spiralis foraminosus through which the fibres of the cochlear nerve pass.
The cochlear canal, about 35 mm long, is arranged spirally around the modiolus and makes two and three-fourths turns. The apical turn begins at the cupula, and the basal turn is continuous behind with the vestibule. The canal increases in diameter from apex to base, and the basal turn bulges into the tympanic cavity as the promontory.
An osseous spiral lamina projects from the surface of the modiolus into the cochlear canal like the thread of a screw. The lamina extends about half-way across the width of the canal and gradually narrows from the basal to the apical turn of cochlea.
The free edge of the lamina splits into a tympamic lip and a vestibular lip separated by a С-shaped sulcus spiralis internus. From the tympanic lip of the lamina a fibrous basilar membrane extends to the crista basilaris on the outer wall of the cochlear canal. Thus the osseous lamina and the basilar membrane together divide the cochlear canal into an upper part, the scala vestibuli (towards the apex), and a lower part, the scala tympani (towards the base).
Both scalae are filled with perilymph and communicate with each other at the apex of the modiolus through an aperture, the helicotrema, where the osseous lamina forms a hook-shaped process. At the basal turn of cochlea the scala tympani is closed by the fusion of the down-turned edge of osseous lamina with the cochlear canal, whereas the scala vestibuli communicates with the anterior wall of the vestibule. Close to the basal turn, the scala tympani presents two features:
(a) Fenestra cochleae, which opens into the tympanic cavity below and behind the promontory, but is closed in the recent state by a trilaminar secondary tympanic membrane. When the perilymphatic fluid is compressed by the inward movement of the foot-plate of the stapes during transmission of sound waves, the fluid is decompressed by the compensatory outward bulging of the secondary tympanic membrane.
(b) Aqueduct of cochlea is a narrow tubular canal which communicates the perilymph with the cerebro-spinal fluid of sub-arachnoid space through the cochlear canaliculus. Some workers suggest the existence of a partition between the perilymph and the CSF. But the electron dense tracer substance, when introduced in the sub-arachnoid space, appears in the perilymph suggesting that the partition presents sieve-like appearance.
The basilar membrane is taut and short (0.2 mm) at the basal turn, but lax and broad (0.4 mm) at the apical turn. The vestibular (Reissner's) membrane is attached close to the vestibular lip of the osseous lamina and extends across the scala vestibuli to the outer wall of the cochlear canal. The triangular area, thus established by the vestibular and basilar membranes and the outer wall of the canal, forms the cochlear duct (scala media).

Vestibule:
The vestibule is a bilaterally compressed, ovoid bony cavity, and intervenes between the cochlea in front and the three semicircular canals behind. Anteriorly, the vestibule communicates with the scala vestibuli and receives the termination of the basal turn of the cochlear duct. Posteriorly and above, the vestibule receives the three semi-circular canals by five openings. The vestibule measures about 5 mm both vertically and antero-posteriorly, and about 3 mm across.
Lateral wall of the vestibule presents the fenestra vestibuli which is closed in the recent state by the base of stapes and annular ligament. The bulging of the ampullated end of the lateral semi-circular canal lies above the fenestra vestibuli. The facial nerve in a bony canal passes laterally above the vestibule and then turns backward in the interval between the fenestra vestibuli and the ampulla of the lateral canal.
Its medial wall presents an opening for the aqueduct of the vestibule which appears beneath the dura mater of the posterior surface of the petrous part of the temporal bone as a tubular blind prolongation of the membranous labyrinth, the saccus and ductus endolymphaticus. The interior of the medial wall of the vestibule is divided by an oblique ridge, the vestibular crest, into a spherical recess below and in front and an elliptical recess above and behind.
The saccule is lodged in the spherical recess which presents foramina for the passage of lower division of the vestibular nerve to the saccule. The utricle is contained in the elliptical recess, whose floor is perforated by the upper division of the vestibular nerve to the utricle, and the ampullae of the superior and lateral semi-circular ducts.
Semi-circular canals:
These bony canals are three in number—anterior (superior), posterior and lateral. Each canal describes two-thirds of a circle, about 15-20 mm long, and communicates by two ends with the upper and posterior part of the vestibule. The medial end of the anterior canal and the upper end of the posterior canal unite to form a crus commune which opens into the medial wall of the vestibule; therefore the semicircular canals open in the vestibule by five openings. One end of each canal presents a dilatation known as the ampulla, which contains the ampullary end of the corresponding semicircular duct.
The anterior canal is convex upward and its antero-lateral end is ampullated; the upward convexity is said to be represented by the arcuate eminence on the anterior surface of the petrous part of temporal bone. The plane of the anterior canal lies at right angle to the long axis of the petrous temporal.
The posterior canal lies parallel with the posterior surface of the petrous temporal, its convexity is directed backward and the ampullated end below. The plane of the anterior canal of one ear is parallel with the posterior canal of the other ear, and vice versa.
The lateral canal becomes horizontal when the head tilts about 30°forward. Its convexity is directed backward, and the antero-lateral end is ampullated.
Membranous labyrinth:
It consists of a closed system of intercommunicating membranous sacs and ducts within the bony labyrinth and is filled with endolymph. The membranous labyrinth presents—the cochlear duct within the bony cochlea, the saccule and utricle within the vestibule, and the three semicircular ducts within the corresponding bony canal (Fig. 14.17).

The semi-circular ducts open into the utricle which joins with the saccule by a Y-shaped utriculo-saccular duct. The saccule in turn communicates in front with the cochlear duct through the ductus reuniens. The utricle, saccule and the semi-circular ducts are suspended from the bony labyrinth by fibro-cellular bands across the perilymphatic space.
The membranous labyrinth possesses two separate functional components—the cochlear duct for hearing (auditory system); the saccule, utricle and semi-circular ducts for balancing (vestibular or labyrinthine system).
The saccule and utricle monitor the static equilibrium and linear acceleration, whereas the semi-circular ducts are concerned with the kinetic equilibrium and angular acceleration. Obviously, the heterogenous functions of the membranous labyrinth are associated with regional structural variations and separate nerve connections.
Cochlear duct (Scala media):
The duct lies along the outer wall of the bony cochlear canal and undergoes a spiral course of two and three- fourths turns. It begins blindly at the cupula and the blind end is known as the lagaena.
The basal turn of the duct enters the vestibule, where it joins with the saccule by a narrow duct, the ductus reuniens. The cochlear duct contains the spiral organ of Corti (peripheral organ of hearing), which rests on the basilar membrane and is covered by the membrana tectoria.
Structure of the cochlear duct (Fig. 14.18):

The cochlear duct is bounded medially by the vestibular (Reissner's) membrane, laterally by the stria vascularis, and below by the tympanic lip of the osseous spiral lamina in the medial part and by the basilar membrane in the lateral part. The basilar membrane is subdivided into a lateral part, zona pectinata, and a medial part, zona arcuata upon which rests the organ of Corti.
The vestibular membrane is made up of an outer layer of flattened epithelial perilymphatic cells, an intervening basal lamina, and the inner endolymphatic surface is lined by a layer of simple squamous epithelium.
The stria vascularis consists of a specialised stratified epithelium which is permeated with a rich plexus of intra-epithelial capillaris. The epithelium is composed of three types of cells— inner dark or chromophil cells, intermediate light or chromophobe cells and outer basal cells. The stria vascularis is the chief source of synthesis of endolymph which closely resembles the intracellular fluid with high K + and low Na + concentration. The positive electrical potential of the endolymph is largely dependent upon oxygenation level of the epithelial cells.
The projecting free edge of the osseous spiral lamina splits into a tympanic lip and a vestibular lip separated by a С-shaped sulcus spiralis internus. The tympanic lip is perforated by numerous foramina for the passage of the branches of the cochlear nerve.
The tractus spiralis fora-minosus at the base of modiolus (bottom of the internal acoustic meatus) transmits the branches of cochlear nerve; the latter pass through the canals of the modiolus and bend successively outwards to reach the attached margin of the osseous lamina, where the spiral canals enlarge and lodge the bipolar spiral ganglion cells.
The basilar membrane stretches from the tympanic lip of the osseous lamina to the crista basilaris which is a conical projection from the outer wall of the cochlear canal. The upper surface of the vestibular lip presents a number of surface elevations, the auditory teeth which are separated by furrows intersected at right angles.
The vestibular lip gives attachment to the membrana tectoria at the tip and to the vestibular membrane somewhat away from the tip.
Spiral organ of corti:
इसमें आंतरिक और बाहरी छड़ (स्तंभ) कोशिकाएं, आंतरिक और बाहरी बाल कोशिकाएं शामिल हैं, जो डीटर और हेन्सेन की कोशिकाओं और मेम्ब्रेन टैक्टोरिया (चित्र। 14.18) का समर्थन करती है।
आंतरिक और बाहरी रॉड कोशिकाएं प्रत्येक एक पंक्ति में व्यवस्थित होती हैं और उनके विस्तारित आधार द्वारा बेसिलर झिल्ली से जुड़ी होती हैं; उनके ऊपरी सिरे झुकते हैं और एक दूसरे से मिलते हैं, एक त्रिकोणीय आंतरिक, कोरटी की सुरंग द्वारा अलग हो जाते हैं। आंतरिक छड़ लगभग 6000 की संख्या में हैं और बेसिलर झिल्ली के साथ लगभग 6O ro का कोण बनाते हैं। बाहरी छड़ें, लगभग 4000 की संख्या में, बेसिलर झिल्ली के साथ लगभग 40 about का कोण बनाती हैं।
आंतरिक बालों की कोशिकाएं आकार में पिरिफॉर्म होती हैं (टाइप I संवेदी कोशिकाएं), लगभग 3500 की संख्या में और आंतरिक छड़ (चित्र 14.19) के औसत दर्जे की तरफ एक पंक्ति में व्यवस्थित होती हैं।

प्रत्येक आंतरिक बाल कोशिका की मुक्त सतह से ऊपर की ओर लगभग 40 से 60 स्टीरियोसिलिया या माइक्रोविली परियोजना; स्टीरियोकिलिया को यू-आकार के तरीके से व्यवस्थित किया जाता है, जिसमें समरूपता को मोडिओलस की ओर निर्देशित किया जाता है।
आंतरिक बाल कोशिका का आधार दो प्रकार के सिनैप्टिक बुटीक के साथ प्रदान किया जाता है। कोक्लेयर तंत्रिका के अभिवाही तंतु बालों की कोशिकाओं के प्लाज्मा झिल्ली के साथ सीधे अन्तर्ग्रथनी संपर्क में आते हैं। कोक्लेयर तंत्रिका के अपवाही तंतुओं के टर्मिनल बुटोन केवल अभिवाही तंतुओं के पार्श्व पहलू के साथ सिनैप्टिक संपर्क बनाते हैं और सीधे बाल कोशिकाओं तक नहीं पहुंचते हैं।
यह बताता है कि अपवाही तंतु अभिवाही तंतुओं के संचरण को नियंत्रित करते हैं। अन्तर्ग्रथनी बैटन के अलावा, आंतरिक बालों की कोशिकाएं आंतरिक फैलेंगल सहायक कोशिकाओं से घिरी होती हैं।
बाहरी बालों की कोशिकाएँ आकार में बेलनाकार होती हैं (टाइप II संवेदी कोशिकाएँ), लगभग 12, 000 से 20, 000 की संख्या में, और बेसल कॉइल में तीन पंक्तियों में, मध्य कुंडली में चार पंक्तियों और एपिकल कॉइल में पाँच पंक्तियों में व्यवस्थित होती हैं। कुल मिलाकर, कोर्टी के सर्पिल अंग में लगभग 23, 500 बाल कोशिकाएं हैं और मनुष्य में कर्णावत तंत्रिका में लगभग बराबर न्यूरॉन्स हैं। प्रत्येक बाहरी हेयर सेल में 80 से 100 स्टीरोकोलिया होते हैं और वे यू या डब्ल्यू-आकार के तरीके से प्रत्येक सेल में व्यवस्थित होते हैं, जिसमें मोडोलियस को निर्देशित किया जाता है।
श्रवण बाल कोशिका कोई किनोसिलियम प्रस्तुत नहीं करती है, लेकिन यू या डब्ल्यू के उत्तल पक्ष पर एक सेंट्रीओल और बेसल शरीर होता है। जब स्टिरियोसिलिया सेंट्रीओल्स की ओर संकुचित होता है, संवेदी बाल कोशिकाएं उत्तेजित होती हैं। बाहरी बालों की कोशिकाओं के आधार के प्लाज्मा झिल्ली कोक्लेयर तंत्रिका के दोनों अभिवाही और अपवाही फाइबर के सिनैप्टिक बुटोन को सीधे प्राप्त करते हैं। यह पता चलता है कि अपवाही प्रभाव से, तंतुमय तंतु बाहरी बालों की कोशिकाओं की उत्तेजना की सीमा को बदल देते हैं।
बाहरी बाल कोशिकाओं की पंक्तियों के बीच Deiter की सहायक कोशिकाएँ हस्तक्षेप करती हैं। वे अपने ठिकानों से बेसिलर झिल्ली तक तय कर रहे हैं, बालों की कोशिकाओं के आधार के चारों ओर समान निवेश बनाते हैं और बालों की कोशिकाओं के बीच जालीदार प्रक्रियाओं के रूप में विस्तार करते हैं।
उत्तरार्द्ध को कई फालन्गेल प्रक्रियाओं के किनारों के तंग जंक्शनों द्वारा एक नाजुक झिल्ली के रूप में बनाया जाता है और बाहरी रॉड कोशिकाओं के सिर से बाहरी बाल कोशिकाओं की बाहरी पंक्ति तक फैली हुई है। जालीदार लामिना बाहरी बालों की कोशिकाओं के मुक्त छोर के लिए गोलाकार छेद प्रस्तुत करता है। जालीदार लैमिना के ऊपरी दो-तिहाई बाहरी बालों की कोशिकाओं के नीचे क्लीयर-जैसे इंटरकंप्युटिंग स्पेस जो कि एक तरल पदार्थ, कॉर्टिल्मफ से भरे होते हैं, से डेयर्स कोशिकाओं से अलग हो जाते हैं।
सहायक हेन्सन की कोशिकाएँ लम्बी स्तंभ कोशिकाएँ होती हैं और बाहरी बालों की कोशिकाओं की बाहरी पाँच या छः पंक्तियों के बाहर होती हैं और क्लॉडियस के क्यूबिकल कोशिकाओं द्वारा बाह्य रूप से अधिक सफल होती हैं; उत्तरार्द्ध कोशिकाओं के साथ निरंतर हैं जो सल्कस स्पिरिलालिस एक्सटर्नलस को लाइन करते हैं।
कोर्टी (आंतरिक सुरंग) की सुरंग के अलावा, सर्पिल अंग बाहरी बालों की कोशिकाओं और हेन्सेन की कोशिकाओं की सबसे बाहरी पंक्ति और बाहरी छड़ी कोशिकाओं और बाहरी बालों की कोशिकाओं के अंतरतम पंक्ति के बीच Nuel के बीच एक बाहरी सुरंग रखता है। सर्पिल अंग के भीतर ये रिक्त स्थान एक दूसरे के साथ संवाद करते हैं और कॉर्टिल्मफ से भरे होते हैं। Cortilymph पेरीलिम्फ जैसा दिखता है, एंडोलिम्फ नहीं; इसमें उच्च Na + और निम्न K + सांद्रता है।
कॉर्टिल्मफ की आयनिक संरचना को बेसिलर झिल्ली द्वारा बनाए रखा जाता है जो आयनों के लिए छलनी के रूप में कार्य करता है जो कि स्केला टिम्पेनी के पेरिलिम्फ से गुजरता है। दूसरी ओर, जालीदार लैमिना एक आयनिक अवरोधक के रूप में कार्य करता है जो एंडोलिम्फ और कॉर्टिल्मफ के बीच विनिमय को रोकता है। उच्च Na + और कम K + कॉर्टिलीमफ की सांद्रता, उत्तेजित करने योग्य बालों की कोशिकाओं के लिए एक सामान्य आयनिक वातावरण बनाती है।
झिल्लीदार टेक्टेरिया एपिडर्मल केरातिन के समान एक एककोशिकीय समर्थक टी-रिच फाइब्रिलेटेड जिलेटिनस संरचना है। यह ओशियस लैमिना के वेस्टिबुलर होंठ से जुड़ा हुआ है और आंतरिक और बाहरी बालों की कोशिकाओं के स्टीरियोकिलिया को कवर करता है। हार्डनेस की झिल्ली अपनी सतह के नीचे एक विशेष क्षेत्र है, जहां बाहरी बालों की कोशिकाओं के स्टीरियोकिलिया के सुझावों को एम्बेडेड किया जाता है। हेन्सन की पट्टी अपने अंडरस्फेस पर एक और क्षेत्र है जहां आंतरिक बालों की कोशिकाओं के स्टीरियोस्किलिया के सुझाव संपर्क में आते हैं।
कोक्लेयर तंत्रिका के परिधीय तंतु आंतरिक और बाहरी बालों की कोशिकाओं के आधार के आसपास मंडराते हैं। अस्थि सर्पिल लैमिना की दो तालिकाओं के बीच से गुजरने के बाद तंत्रिका तंतु मोडिओलस में प्रवेश करते हैं। मोडिओलस के भीतर प्रत्येक फाइबर द्विध्रुवीय सर्पिल नाड़ीग्रन्थि कोशिकाओं की परिधीय प्रक्रिया के रूप में समाप्त होता है; नाड़ीग्रन्थि कोशिकाओं की केंद्रीय प्रक्रियाएं आंतरिक ध्वनिक मांस के तल में ट्रैक्टस स्पाइरलिस फॉर्मिनोस से गुजरती हैं और कोक्लेयर तंत्रिका के ट्रंक को बनाने के लिए एकजुट होती हैं।
यह अनुमान है कि दस सर्पिल नाड़ीग्रन्थि कोशिकाएं प्रत्येक आंतरिक बाल कोशिका से जुड़ी होती हैं, जबकि प्रत्येक नाड़ीग्रन्थि कोशिका दस से अधिक बाहरी बाल कोशिकाओं की आपूर्ति करती है। आंतरिक बालों की कोशिकाएं रेटिना के शंकु के अनुरूप होती हैं और ध्वनि तरंगों के ठीक भेदभाव के लिए जिम्मेदार होती हैं, जबकि बाहरी बाल कोशिकाएं नजदीकी दहलीज घटनाओं का पता लगाने में अधिक शामिल होती हैं। कोक्लीया के बेसल मोड़ में सर्पिल अंग उच्च-पिच ध्वनि के साथ संबंधित है, कि कम-पिच वाली ध्वनि के साथ एपिक मोड़ में।
Saccule और utricle (चित्र। 14.17):
बोनी वेस्टिबुल के भीतर थैली और यूरीकल दोनों ही सीमित हैं। उनमें से प्रत्येक के पास विशेष इंद्रिय अंग होते हैं, मैक्युला, जो संवेदनशील बाल कोशिकाओं के गुरुत्वाकर्षण रिसेप्टर्स द्वारा स्थिर संतुलन और रैखिक त्वरण को नियंत्रित करता है।
सेक्युल:
यह एक गोलाकार झिल्लीदार थैली है, जो वेस्टिब्यूल के गोलाकार अवकाश में स्थित है। Saccule डक्टस रीयूनियन के माध्यम से कर्णावत वाहिनी के सामने संचार करता है। यह वाई-आकार के यूट्रिकुलोसैक्युलर डक्ट द्वारा यूरीकल के पीछे जुड़ा हुआ है जो वेस्टिब्यूल के एक्वाडक्ट से गुजरने के बाद ड्यूरा मेटर के नीचे एक अंधा सैकस एंडोलिम्फैटिकस बनाता है; एंडोलिमिक उपकला कोशिकाओं को अणु द्वारा अवशोषित कर लेता है। मैक्युला, लगभग 2 मिमी 3 मिमी आकार का, थैली की मध्य दीवार में स्थित है और वेस्टिबुलर तंत्रिका के निचले विभाजन द्वारा संक्रमित है।
यूट्रिकल:
यह आकार में तिरछा है, अण्डाकार अवकाश में थैली के ऊपर और पीछे स्थित है। इसके बाद के संस्करण और बाद में, यूट्रिकल तीन अर्धवृत्ताकार नलिकाओं को पांच छिद्रों द्वारा प्राप्त करता है। कभी-कभी utricle के फर्श और saccule की छत को बारीकी से अनुमानित किया जाता है। यूट्रिकल का मैक्युला फर्श और पूर्वकाल की दीवार के साथ स्थित है; यह वेस्टिबुलर तंत्रिका के ऊपरी विभाजन द्वारा संक्रमित है।
मैक्युला की संरचना:
मूल रूप से झिल्लीदार भूलभुलैया के वेस्टिबुलर घटक की दीवार में तीन परतें होती हैं (छवि 14.20):
(ए) पेरिलिम्पैटिक कोशिकाओं द्वारा पंक्तिबद्ध एक बाहरी परत;
(b) ट्युनिका प्रोप्रिया की मध्य परत;
(c) आंतरिक परत में उपकला कोशिकाओं की एक पंक्ति होती है।

मैक्यूला में, आंतरिक परत बालों की कोशिकाओं को प्रस्तुत करती है, कोशिकाओं का समर्थन करती है और जिलेटिनस द्रव्यमान की एक पट्टिका द्वारा कवर की जाती है जिसे ओटोलिथिक झिल्ली के रूप में जाना जाता है जो कैल्शियम क्रिस्टल, कैल्साइट के साथ गर्भवती होती है। ओटोलिथिक झिल्ली में स्टेरोकिलिया और किनोसिलियम परियोजना।
बालों की कोशिकाएँ संवेदी कोशिकाएँ होती हैं और प्रत्येक को 70-80 स्टिरियोसिलिया और एक किनोसिलियम के साथ मुक्त सतह पर प्रदान किया जाता है। स्टिरियोसिलिया संशोधित माइक्रोविल्ली हैं और किनोसिलिया संशोधित सिलिया हैं। कोशिका के एक सिरे पर एक लंबी कीनोकोइलियम होती है, और लंबाई के घटते क्रम का स्टाइनोसिलिया किनोसिलियम से दूर एक पंक्ति में स्थित होता है। यह अभिविन्यास मोर्फोलॉजिक ध्रुवीकरण पैदा करता है। जब स्टीरोकोलिया किनोसिलियम की ओर विचलित हो जाता है, तो बाल कोशिका को विध्रुवण (उत्तेजना) में ले जाता है; जब वे विपरीत दिशा में विचलन करते हैं, तो बाल कोशिका हाइपरपोलराइज्ड (अवरोध) होती है। प्रत्येक बाल कोशिका की सतह पर किनोकिलिया के स्थान एक विभाजन रेखा के संबंध में सैक्यूल और यूरीकल के मैक्युले में भिन्न होते हैं जो मैक्युला के मध्य क्षेत्र के माध्यम से खींचा जाता है। Saccule में kinocilia को बिदाई लाइन से दूर निर्देशित किया जाता है, जबकि यूट्रिकल में वे लाइन की ओर स्थित होते हैं (चित्र 14.21)।

आकार और प्रकृति की प्रकृति के अनुसार, बाल कोशिकाएं दो प्रकार की होती हैं:
(ए) प्रकार मैं बालों की कोशिकाओं को एक गोल आधार के साथ फ्लास्क के आकार का होता है, और एक श्लेष्म की तरह चक्करदार तंत्रिका अंत द्वारा संलग्न किया जाता है। वेस्टिबुलर अपवाही तंतु अभिवाही तंत्रिका अंत के साथ सिनाप्सेस बनाते हैं, और इस तरह निषेध द्वारा संचरण संकेतों को विनियमित करते हैं।
(b) टाइप II हेयर सेल्स लम्बी स्तंभकार हैं, और कई अभिवाही और अपवाही सिनैप्टिक बॉटन बालों की कोशिकाओं के साथ सीधा संपर्क बनाते हैं। इस प्रकार निरोधात्मक अपवाही तंतु सीधे बालों की कोशिकाओं के उत्तेजना की दहलीज को नियंत्रित करते हैं।
सहायक कोशिकाएं स्तंभित होती हैं, एक बेसल लामिना पर आराम करती हैं और ओटोलिथिक झिल्ली में पेशी से मुक्त सतह पर माइक्रोविली पेश करती हैं।
Saccule और utricle के कार्य:
(ए) जब कैल्शियम क्रिस्टल के साथ ओटोलिथिक झिल्ली सिर की गति से नीचे की ओर आती है, तो यह संतरी बालों की कोशिकाओं पर गुरुत्वाकर्षण खींचती है और शरीर के संबंध में सिर की स्थिति की जानकारी देती है।
(b) एक ही तरफ सिर के पार्श्व झुकाव के दौरान थैली को उत्तेजित किया जाता है। सिर को या तो आगे या पीछे की ओर झुकाए जाने पर उत्कीर्णन उत्तेजित होता है।
(c) यूट्रिकल और संभवतः सैक्यूल एक सीधी रेखा में सिर के आंदोलन के दौरान रैखिक त्वरण का जवाब देते हैं- आगे, पीछे, ऊपर या नीचे। इन सेंसरों की विफलता बताती है कि क्यों एक हवाई जहाज का पायलट, जब एक छोटे विमान में बादलों के माध्यम से बिना उपकरणों के उड़ान भरता है, तो बिना एहसास किए उल्टा उड़ सकता है।
अर्धवृत्ताकार नलिकाएं:
तीन अर्धवृत्ताकार नलिकाएं संबंधित बोनी नहरों के भीतर समाहित हैं। प्रत्येक नलिका नहर की बाहरी दीवार के साथ गुजरती है और बोनी नहर के व्यास का लगभग 1 / 4th है। Ampulla के भीतर नलिका कमजोर पड़ जाती है और crista ampullaris को प्रस्तुत करती है जो वाहिनी के आंतरिक भाग में प्रोजेक्ट करती है। अर्ध-वृत्ताकार नलिकाएं पांच छिद्रों द्वारा खोलती हैं (अंजीर। 14.17, 14.22)।

Crista ampullaris की संरचना:
Ampulla के उपकला कोशिकाओं में संवेदी बाल कोशिकाएं, स्तंभ सहायक कोशिकाएं होती हैं और एक शंकु के आकार का जलीय द्रव्यमान, कपुला, जिसमें बालों की कोशिकाओं के स्टिरियोसिलिया और किनोसिलिया शामिल होते हैं। बाल कोशिकाओं के आधार को वेस्टिबुलर तंत्रिका (छवि 14.23) द्वारा जन्म दिया जाता है।

बालों की कोशिकाओं के प्रकार और स्टिरियोसिलिया और किनोसिलिया के उन्मुखीकरण मैक्युले के समान हैं। पार्श्व अर्धवृत्ताकार वाहिनी के ampulla में, बालों की कोशिकाओं के किनोसीलिया को यूट्रिकल की ओर निर्देशित किया जाता है, जबकि पूर्वकाल और पीछे के नलिकाओं के ampulla में वे यूरीकल से दूर निर्देशित होते हैं।
अर्धवृत्ताकार नलिकाओं के कार्य:
(ए) crista ampullaris का कपूला एक झूले-दरवाजे की तरह काम करता है और एंडोलिम्फ के आंदोलनों द्वारा एक तरफ से विक्षेपित होता है। यह रोटरी रिसेप्टर के रूप में कार्य करता है और सिर के रोटेशन के दौरान कोणीय त्वरण की निगरानी करता है और एक ही समय में शरीर की मुद्रा को सीधा रखता है।
(b) कीनो-सिलियम के प्रति स्टिरियोसिलिया का संपीडन बाल कोशिकाओं की उत्तेजना पैदा करता है, और उनके विघटन से अवरोध उत्पन्न होता है। इसलिए, पार्श्व (क्षैतिज) वाहिनी को प्रेरित किया जाता है, जब कपुला को बर्तन की ओर विक्षेपित किया जाता है, जबकि ऊर्ध्वाधर नलिकाओं (पूर्वकाल और पीछे) को उत्तेजित किया जाता है, जब कपुला को बर्तन से दूर कर दिया जाता है।
(c) किसी विशेष नहर के बालों की कोशिकाओं को उत्तेजित करना उस विमान पर निर्भर करता है जिसमें घुमाव होता है। नहर सबसे प्रभावी रूप से अपने स्वयं के विकर्ण विमान में रोटेशन द्वारा प्रेरित होती है।
एक ऊर्ध्वाधर अक्ष के चारों ओर सिर का रोटरी आंदोलन पार्श्व अर्धवृत्ताकार नलिकाओं को उत्तेजित करता है (छवि 14.24)।

एक घड़ी-वार घुमाव दाहिने वाहिनी को उत्तेजित करता है, और एक एंटीलॉक-वार घुमाव बाएं वाहिनी को उत्तेजित करता है। जब एक घड़ी-वार घुमाव शुरू होता है, तो एंडोलिम्फ की जड़ता दाईं कपुला को यूट्रिकल की ओर और लेफ्ट कपुला को यूरीकल से दूर कर देती है। आखिरकार दाहिनी नलिका उत्तेजित होती है, और बाईं वाहिनी बाधित होती है। जब एक ही गति के साथ रोटरी आंदोलन जारी रहता है और एंडोलिम्फ की जड़ता दूर हो जाती है, तो कपुले अपने आराम करने वाले पदों को पुनः प्राप्त करते हैं। आंदोलन की समाप्ति पर, तरल पदार्थ की गति विपरीत दिशा में कपुले के झुकने का कारण बनती है। यही कारण है कि लंबे समय तक रोटेशन के दौरान कोई सनसनी नहीं होती है, लेकिन विपरीत दिशा में मुड़ने की सनसनी तब महसूस होती है जब रोटेशन अचानक बंद हो जाता है।
वेस्टिबुलो-कॉक्लियर तंत्रिका:
यह VUIth कपाल तंत्रिका है और उन दोनों के बीच में हस्तक्षेप करने वाले तंत्रिका मध्यस्थ के साथ चेहरे और तंत्रिका के पीछे के पोन्स और मज्जा पुलाव के बीच जंक्शन से जुड़ा हुआ है। वेस्टिबुलो-कोक्लेयर तंत्रिका में दो घटक होते हैं- संतुलन के लिए वेस्टिबुलर तंत्रिका, और सुनवाई के लिए कोक्लेयर तंत्रिका। मस्तिष्क के तने के करीब, वेस्टिबुलर तंत्रिका कोक्लेयर तंत्रिका को वेंट्रो-मेडियल होता है।
वेस्टिबुलर तंत्रिका:
इसमें लगभग 19, 000 तंत्रिका तंतु शामिल हैं और निम्नलिखित कार्यात्मक घटक प्रस्तुत करते हैं:
(ए) saccule और utricle के maculae के संवेदनशील बालों की कोशिकाओं से स्थिर संतुलन के लिए विशेष प्रोप्रियोसेप्टिव फाइबर;
(बी) तीन अर्धवृत्ताकार नलिकाओं के ampullary जंगलों के बालों की कोशिकाओं से गतिज संतुलन के लिए विशेष प्रोप्रियोसेप्टिव फाइबर;
(c) ओलिवो-कोक्लेयर बंडल के कुछ अपवाही तंतु टाइप II बाल कोशिकाओं तक सीधे पहुँचते हैं और प्रकार मैं बाल कोशिकाएँ टर्मिनल अभिवाही बुटीक के साथ सिनैप्स बनाकर। जिससे अपवाही तंतु अवरोध द्वारा अंश की डिग्री को नियंत्रित करते हैं।
उत्पत्ति के सेल :
1. स्थैतिक और गतिज संतुलन दोनों के लिए अभिवाही तंतु द्विध्रुवी न्यूरॉन्स से उत्पत्ति लेते हैं
वेस्टिबुलर गैंग्लियन (स्कार्पा का गैंग्लियन) जो आंतरिक ध्वनिक मांस के तल पर वेस्टिबुलर तंत्रिका के ट्रंक में स्थित है।
2. अपसारी और गर्भ-पार्श्व दोनों पक्षों से, तंतुओं के बेहतर और सहायक ऑलिवरी नाभिक से अपवाही तंतु उत्पन्न होते हैं। कुछ अपवर्तक तंतु वेस्टिबुलर से कोक्लेयर तंत्रिकाओं तक विस्तार करते हैं जो वेस्टिबुलो-कोक्लेयर एनास्टोमोसिस (ऊर्ट बंडल) द्वारा होता है।
वेस्टिबुलर तंत्रिका का परिधीय वितरण:
आंतरिक मांस के तल पर द्विध्रुवी नाड़ीग्रन्थि कोशिकाओं की परिधीय प्रक्रियाएं ऊपरी, निचले और पीछे की शाखाओं में विभाजित होती हैं। ऊपरी शाखाएं श्रेष्ठ वेस्टिबुलर क्षेत्र के अग्रभाग से होकर गुजरती हैं, जो पूर्वकाल और पार्श्व अर्धवृत्ताकार नलिकाओं के उदरस्थ और ampullary crests के मैक्युला तक पहुंचती हैं। निचली शाखाएं अवर वेस्टिबुलर क्षेत्र की फोरामिना के माध्यम से थैली के मैक्युला तक पहुंचती हैं; पीछे की शाखा फोरमैन सिंगुलारे के माध्यम से प्रवेश करती है और पश्च अर्धवृत्ताकार वाहिनी के ampullary शिखा में समाप्त होती है।
आंतरिक मांस के तल को हड्डी की एक प्लेट द्वारा सीमित किया जाता है जो ऊपरी और निचले क्षेत्रों में एक क्षैतिज शिखा द्वारा उप-विभाजित होता है। ऊपरी क्षेत्र के पीछे का हिस्सा बेहतर वेस्टिबुलर क्षेत्र को प्रस्तुत करता है, और निचले क्षेत्र का अवर अवर वेस्टिबुलर क्षेत्र द्वारा कब्जा कर लिया जाता है। ऊपरी क्षेत्र का पूर्वकाल हिस्सा चेहरे की तंत्रिका के लिए एक उद्घाटन प्रस्तुत करता है, और निचले क्षेत्र के कर्णावर्त तंत्रिका के तंतुओं के संचरण के लिए ट्रैक्टस स्पिरिलालिस फॉरमिनोसस प्रदर्शित करता है।
वेस्टिबुलर तंत्रिका के केंद्रीय कनेक्शन:
वेस्टिबुलर तंत्रिका के ट्रंक से द्विध्रुवी नाड़ीग्रन्थि कोशिकाओं की केंद्रीय प्रक्रियाएं जो कर्णावत तंत्रिका के वेंट्रोमेडियल भाग पर कब्जा करने के लिए सर्पिल फैशन में औसत दर्जे से गुजरती हैं।
पोन्स की निचली सीमा तक पहुँचने पर, वेस्टिबुलर तंत्रिका अवर सेरेबेलर पेडुंक्ल से औसत दर्जे का गुजरता है, आरोही और अवरोही शाखाओं में विभाजित होता है और वेस्टिबुलर नाभिक के चार समूहों (श्रेष्ठ, अवर, औसत दर्जे का और पार्श्व) के साथ सिंकैप्स बनाता है; कुछ फाइबर, हालांकि, सीधे जुकोटा-रेस्टीफॉर्म बॉडी के माध्यम से सेरिबैलम के फ्लोकुलो-नोड्यूलर लोब तक सीधे पहुंचते हैं।
कर्णावत तंत्रिका:
इसमें लगभग 23, 500 फाइबर होते हैं और निम्नलिखित कार्यात्मक घटक होते हैं:
(ए) अधिकांश फाइबर कोर्टी के अंग के बाल कोशिकाओं (फोनोसेप्टर्स) से सुनने के लिए विशेष दैहिक सनसनी को व्यक्त करते हैं;
(b) ओलिवो-कोक्लेयर बंडल के कुछ अपवाही तंतु सीधे या परोक्ष रूप से एक्जिट-बाइल की डिग्री को संशोधित करने और बालों की कोशिकाओं को तेज आवाज़ से बचाने के लिए बालों की कोशिकाओं तक पहुंचते हैं।
उत्पत्ति के सेल:
अभिवाही तंतु द्विध्रुवीय सर्पिल नाड़ीग्रन्थि कोशिकाओं से उत्पन्न होते हैं जो मोडिओलस के सर्पिल नहर के भीतर स्थित होते हैं।
परिधीय वितरण:
द्विध्रुवीय सर्पिल न्यूरॉन्स की परिधीय प्रक्रियाएं असमस लामिना के संलग्न मार्जिन तक पहुंचने के लिए क्रमिक रूप से बाहर की ओर झुकती हैं, और ऑसियस लामिना की दो तालिकाओं के बीच से गुजरने के बाद तंत्रिका तंतु आंतरिक और बाहरी बालों की कोशिकाओं के आधारों के आसपास मंडराते हैं। प्रत्येक आंतरिक बालों की कोशिका को लगभग दस सर्पिल नाड़ीग्रन्थि कोशिकाओं से कनेक्शन प्राप्त होता है, जबकि एक नाड़ीग्रन्थि कोशिका दस से अधिक बाहरी बाल कोशिकाओं से जुड़ी होती है।
केंद्रक कनेक्शन:
नाड़ीग्रन्थि कोशिकाओं की केंद्रीय प्रक्रियाएं आंतरिक ध्वनिक मांस के तल में ट्रैक्टस स्पाइरलिस फॉरेमोनस से गुजरती हैं और कोक्लेयर तंत्रिका के ट्रंक बनाने के लिए एकजुट होती हैं।
पोंस की निचली सीमा पर कोक्लेयर तंत्रिका के तंतु अवर सेरिबैलर पेडुंकल को पार्श्व पास करते हैं और उदर और पृष्ठीय कोक्लेयर नाभिक के साथ सिनैप्स बनाते हैं। (आगे के कनेक्शन के लिए, लेखक की पुस्तक "न्यूरोनेटोमी की अनिवार्यता" मस्तिष्क स्टेम के तहत परामर्श करें)।